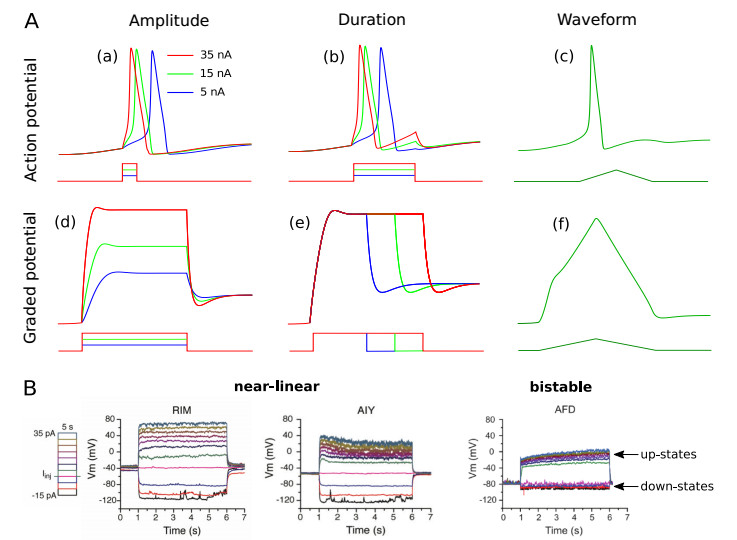
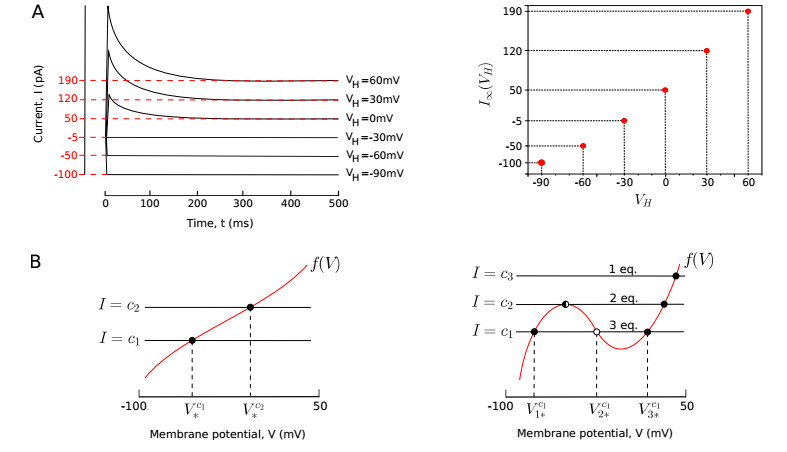
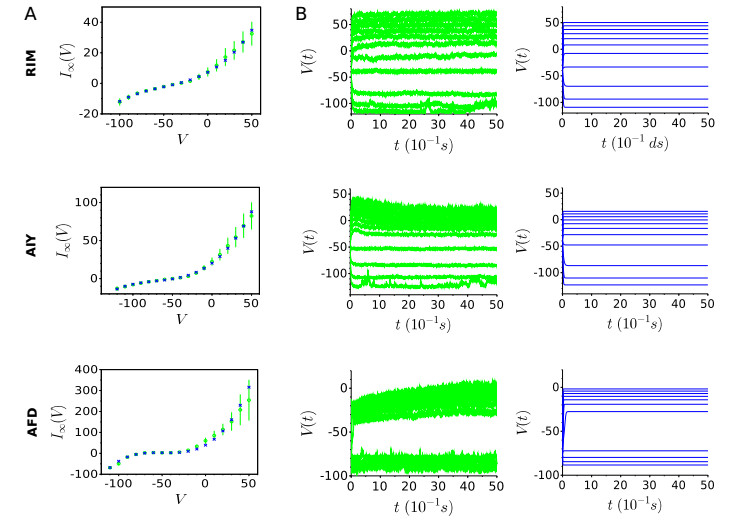
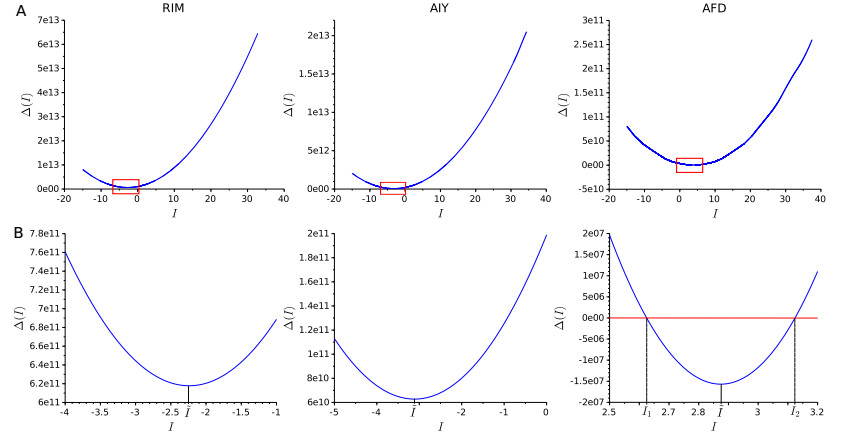
A central goal of neuroscience is to understand the way nervous systems work to produce behavior. Experimental measurements in freely moving animals (e.g. in the C. elegans worm) suggest that ON- and OFF-states in non-spiking nervous tissues underlie many physiological behaviors. Such states are defined by the collective activity of non-spiking neurons with correlated up- and down-states of their membrane potentials. How these network states emerge from the intrinsic neuron dynamics and their couplings remains unclear. In this paper, we develop a rigorous mathematical framework for better understanding their emergence. To that end, we use a recent simple phenomenological model capable of reproducing the experimental behavior of non-spiking neurons. The analysis of the stationary points and the bifurcation dynamics of this model are performed. Then, we give mathematical conditions to monitor the impact of network activity on intrinsic neuron properties. From then on, we highlight that ON- and OFF-states in non-spiking coupled neurons could be a consequence of bistable synaptic inputs, and not of intrinsic neuron dynamics. In other words, the apparent up- and down-states in the neuron's bimodal voltage distribution do not necessarily result from an intrinsic bistability of the cell. Rather, these states could be driven by bistable presynaptic neurons, ubiquitous in non-spiking nervous tissues, which dictate their behaviors to their postsynaptic ones.
Citation: Loïs Naudin. Biological emergent properties in non-spiking neural networks[J]. AIMS Mathematics, 2022, 7(10): 19415-19439. doi: 10.3934/math.20221066
A central goal of neuroscience is to understand the way nervous systems work to produce behavior. Experimental measurements in freely moving animals (e.g. in the C. elegans worm) suggest that ON- and OFF-states in non-spiking nervous tissues underlie many physiological behaviors. Such states are defined by the collective activity of non-spiking neurons with correlated up- and down-states of their membrane potentials. How these network states emerge from the intrinsic neuron dynamics and their couplings remains unclear. In this paper, we develop a rigorous mathematical framework for better understanding their emergence. To that end, we use a recent simple phenomenological model capable of reproducing the experimental behavior of non-spiking neurons. The analysis of the stationary points and the bifurcation dynamics of this model are performed. Then, we give mathematical conditions to monitor the impact of network activity on intrinsic neuron properties. From then on, we highlight that ON- and OFF-states in non-spiking coupled neurons could be a consequence of bistable synaptic inputs, and not of intrinsic neuron dynamics. In other words, the apparent up- and down-states in the neuron's bimodal voltage distribution do not necessarily result from an intrinsic bistability of the cell. Rather, these states could be driven by bistable presynaptic neurons, ubiquitous in non-spiking nervous tissues, which dictate their behaviors to their postsynaptic ones.

| [1] |
H. Markram, E. Muller, S. Ramaswamy, M. W. Reimann, M. Abdellah, C. A. Sanchez, et al., Reconstruction and simulation of neocortical microcircuitry, Cell, 163 (2015), 456–492. https://doi.org/10.1016/j.cell.2015.09.029 doi: 10.1016/j.cell.2015.09.029

|
| [2] |
F. Giovannini, B. Knauer, M. Yoshida, L. Buhry, The can-in network: A biologically inspired model for self-sustained theta oscillations and memory maintenance in the hippocampus, Hippocampus, 27 (2017), 450–463. https://doi.org/10.1002/hipo.22704 doi: 10.1002/hipo.22704
|
| [3] |
F. Cavarretta, G. Naldi, Mathematical study of a nonlinear neuron model with active dendrites, AIMS Mathematics, 4 (2019), 831–846. https://doi.org/10.3934/math.2019.3.831 doi: 10.3934/math.2019.3.831
|
| [4] |
H. Lin, C. Wang, Y. Sun, W. Yao, Firing multistability in a locally active memristive neuron model, Nonlinear Dynamics, 100 (2020), 3667–3683. https://doi.org/10.1007/s11071-020-05687-3 doi: 10.1007/s11071-020-05687-3
|
| [5] |
H. Lin, C. Wang, Q. Deng, C. Xu, Z. Deng, C. Zhou, Review on chaotic dynamics of memristive neuron and neural network, Nonlinear Dynamics, 106 (2021), 959–973. https://doi.org/10.1007/s11071-021-06853-x doi: 10.1007/s11071-021-06853-x
|
| [6] |
H. Lin, C. Wang, C. Chen, Y. Sun, C. Zhou, C. Xu, et al., Neural bursting and synchronization emulated by neural networks and circuits, IEEE T. Circuits-I, 68 (2021), 3397–3410. https://doi.org/10.1109/TCSI.2021.3081150 doi: 10.1109/TCSI.2021.3081150
|
| [7] |
S. R. Lockery, M. B. Goodman, S. Faumont, First report of action potentials in a c. elegans neuron is premature, Nat. Neurosci., 12 (2009), 365–366. https://doi.org/10.1038/nn0409-365 doi: 10.1038/nn0409-365
|
| [8] |
G. D. Field, E. J. Chichilnisky, Information processing in the primate retina: circuitry and coding. Annu. Rev. Neurosci., 30 (2007), 1–30. https://doi.org/10.1146/annurev.neuro.30.051606.094252 doi: 10.1146/annurev.neuro.30.051606.094252
|
| [9] |
M. B. Goodman, D. H. Hall, L. Avery, S. R. Lockery, Active currents regulate sensitivity and dynamic range in c. elegans neurons, Neuron, 20 (1998), 763–772. https://doi.org/10.1016/S0896-6273(00)81014-4 doi: 10.1016/S0896-6273(00)81014-4
|
| [10] | A. Roberts, B. M. H. Bush, Neurones without impulses: their significance for vertebrate and invertebrate nervous systems, volume 6, Cambridge University Press, 1981. |
| [11] |
R. E. Davis, A. O. Stretton, Signaling properties of ascaris motorneurons: graded active responses, graded synaptic transmission, and tonic transmitter release, J. Neurosci., 9 (1989), 415–425. https://doi.org/10.1523/JNEUROSCI.09-02-00415.1989 doi: 10.1523/JNEUROSCI.09-02-00415.1989
|
| [12] |
S. Usui, A. Ishihaiza, Y. Kamiyama, H. Ishii, Ionic current model of bipolar cells in the lower vertebrate retina, Vision res., 36 (1996), 4069–4076. https://doi.org/10.1016/S0042-6989(96)00179-4 doi: 10.1016/S0042-6989(96)00179-4
|
| [13] |
D. E. Kourennyi, X. D. Liu, J. Hart, F. Mahmud, W. H. Baldridge, S. Barnes, Reciprocal modulation of calcium dynamics at rod and cone photoreceptor synapses by nitric oxide, J. Neurophysiol., 92 (2004), 477–483. https://doi.org/10.1152/jn.00606.2003 doi: 10.1152/jn.00606.2003
|
| [14] |
R. Publio, R. F. Oliveira, A. C. Roque, A realistic model of rod photoreceptor for use in a retina network model, Neurocomputing, 69 (2006), 1020–1024. https://doi.org/10.1016/j.neucom.2005.12.037 doi: 10.1016/j.neucom.2005.12.037
|
| [15] |
L. Naudin, N. Corson, M. A. Aziz-Alaoui, J. L. Jimenez Laredo, T. Démare, On the modeling of the three types of non-spiking neurons of the caenorhabditis elegans, Int. J. Neural Syst., 31 (2021), 2050063. https://doi.org/10.1142/S012906572050063X doi: 10.1142/S012906572050063X
|
| [16] |
M. Nicoletti, A. Loppini, L. Chiodo, V. Folli, G. Ruocco, S. Filippi, Biophysical modeling of c. elegans neurons: Single ion currents and whole-cell dynamics of awcon and rmd, PloS one, 14 (2019), e0218738. https://doi.org/10.1371/journal.pone.0218738 doi: 10.1371/journal.pone.0218738
|
| [17] |
L. Naudin, J. L. Jiménez Laredo, Q. Liu, N. Corson, Systematic generation of biophysically detailed models with generalization capability for non-spiking neurons, PloS one, 17 (2022), e0268380. https://doi.org/10.1371/journal.pone.0268380 doi: 10.1371/journal.pone.0268380
|
| [18] |
J. L. Jiménez Laredo, L. Naudin, N. Corson, C. M. Fernandes, A methodology for determining ion channels from membrane potential neuronal recordings, Applications of Evolutionary Computation, (2022), 15–29. https://doi.org/10.1007/978-3-031-02462-7_2 doi: 10.1007/978-3-031-02462-7_2
|
| [19] |
J. G. White, E. Southgate, J. N. Thomson, S. Brenner, The structure of the nervous system of the nematode caenorhabditis elegans, Philos Trans R Soc Lond B Biol Sci, 314 (1986), 1–340. https://doi.org/10.1098/rstb.1986.0056 doi: 10.1098/rstb.1986.0056
|
| [20] |
L. R. Varshney, B. L. Chen, E. Paniagua, D. H. Hall, D. B. Chklovskii, Structural properties of the caenorhabditis elegans neuronal network, PLoS Comput. Biol., 7 (2011), e1001066. https://doi.org/10.1371/journal.pcbi.1001066 doi: 10.1371/journal.pcbi.1001066
|
| [21] |
S. J. Cook, T. A. Jarrell, C. A. Brittin, Y. Wang, A. E. Bloniarz, M. A. Yakovlev, et al., Whole-animal connectomes of both caenorhabditis elegans sexes, Nature, 571 (2019), 63–71. https://doi.org/10.1038/s41586-019-1352-7 doi: 10.1038/s41586-019-1352-7
|
| [22] |
S. H. Chalasani, N. Chronis, M. Tsunozaki, J. M. Gray, D. Ramot, M. B. Goodman, et al., Dissecting a circuit for olfactory behaviour in caenorhabditis elegans, Nature, 450 (2007), 63–70. https://doi.org/10.1038/nature06292 doi: 10.1038/nature06292
|
| [23] |
P. Liu, B. Chen, Z. W. Wang, Gabaergic motor neurons bias locomotor decision-making in c. elegans, Nat. commun., 11 (2020), 1–19. https://doi.org/10.1038/s41467-020-18893-9 doi: 10.1038/s41467-020-18893-9
|
| [24] |
K. T. Quach, S. H. Chalasani, Flexible reprogramming of pristionchus pacificus motivation for attacking caenorhabditis elegans in predator-prey competition, Curr. Biol., 32 (2022), 1675–1688. https://doi.org/10.1101/2021.03.09.434602 doi: 10.1101/2021.03.09.434602
|
| [25] |
P. Heyward, M. Ennis, A. Keller, M. T. Shipley, Membrane bistability in olfactory bulb mitral cells, J. Neurosci., 21 (2001), 5311–5320. https://doi.org/10.1523/JNEUROSCI.21-14-05311.2001 doi: 10.1523/JNEUROSCI.21-14-05311.2001
|
| [26] |
Y. Loewenstein, S. Mahon, P. Chadderton, K. Kitamura, H. Sompolinsky, Y. Yarom, et al., Bistability of cerebellar purkinje cells modulated by sensory stimulation, Nat. Neurosci., 8 (2005), 202–211. https://doi.org/10.1038/nn1393 doi: 10.1038/nn1393
|
| [27] |
S. W. Hughes, D. W. Cope, T. I. Tóth, S. R. Williams, V. Crunelli, All thalamocortical neurones possess a t-type ca2+ 'window' current that enables the expression of bistability-mediated activities, The Journal of physiology, 517 (1999), 805–815. https://doi.org/10.1111/j.1469-7793.1999.0805s.x doi: 10.1111/j.1469-7793.1999.0805s.x
|
| [28] |
T. Schrödel, R. Prevedel, K. Aumayr, M. Zimmer, A. Vaziri, Brain-wide 3d imaging of neuronal activity in caenorhabditis elegans with sculpted light, Nat. methods, 10 (2013), 1013–1020. https://doi.org/10.1038/nmeth.2637 doi: 10.1038/nmeth.2637
|
| [29] |
R. Prevedel, Y.-G. Yoon, M. Hoffmann, N. Pak, G. Wetzstein, S. Kato, et al., Simultaneous whole-animal 3d imaging of neuronal activity using light-field microscopy, Nat. methods, 11 (2014), 727–730. https://doi.org/10.1038/nmeth.2964 doi: 10.1038/nmeth.2964
|
| [30] |
A. Gordus, N. Pokala, S. Levy, S. W. Flavell, C. I. Bargmann, Feedback from network states generates variability in a probabilistic olfactory circuit, Cell, 161 (2015), 215–227. https://doi.org/10.1016/j.cell.2015.02.018 doi: 10.1016/j.cell.2015.02.018
|
| [31] | E. M. Izhikevich, Dynamical systems in neuroscience, MIT press, 2007. https://doi.org/10.7551/mitpress/2526.001.0001 |
| [32] |
L. Naudin, J. L. Jiménez Laredo, N. Corson, A simple model of non-spiking neurons, Neural Comput., 34 (2022). https://doi.org/10.1162/neco_a_01531 doi: 10.1162/neco_a_01531
|
| [33] |
Q. Liu, P. B. Kidd, M. Dobosiewicz, C. I. Bargmann, C. elegans awa olfactory neurons fire calcium-mediated all-or-none action potentials, Cell, 175 (2018), 57–70. https://doi.org/10.1016/j.cell.2018.08.018 doi: 10.1016/j.cell.2018.08.018
|
| [34] | L. Naudin, N. Corson, M. A. A. Alaoui, A generic conductance-based model of non-spiking caenorhabditis elegans neurons and its mathematical analysis, 2021. |
| [35] |
R. Storn, K. Price, Differential evolution–a simple and efficient heuristic for global optimization over continuous spaces, J. Global Optim., 11 (1997), 341–359. https://doi.org/10.1023/A:1008202821328 doi: 10.1023/A:1008202821328
|
| [36] |
R. Sarpeshkar, Analog versus digital: extrapolating from electronics to neurobiology, Neural Comput., 10 (1998), 1601–1638. https://doi.org/10.1162/089976698300017052 doi: 10.1162/089976698300017052
|
| [37] |
J. G. White, E. Southgate, J. N. Thomson, S. Brenner, The structure of the nervous system of the nematode caenorhabditis elegans, Philos Trans R Soc Lond B Biol Sci, 314 (1986), 1–340. https://doi.org/10.1098/rstb.1986.0056 doi: 10.1098/rstb.1986.0056
|
| [38] |
S. L. Mclntire, E. Jorgensen, J. Kaplan, H. R. Horvitz, The gabaergic nervous system of caenorhabditis elegans, Nature, 364 (1993), 337–341. https://doi.org/10.1038/364337a0 doi: 10.1038/364337a0
|
| [39] |
E. Serrano-Saiz, R. J. Poole, T. Felton, F. Zhang, E. D. De La Cruz, O. Hobert, Modular control of glutamatergic neuronal identity in c. elegans by distinct homeodomain proteins, Cell, 155 (2013), 659–673. https://doi.org/10.1016/j.cell.2013.09.052 doi: 10.1016/j.cell.2013.09.052
|
| [40] |
L. Pereira, P. Kratsios, E. Serrano-Saiz, H. Sheftel, A. E. Mayo, D. H. Hall, et al., A cellular and regulatory map of the cholinergic nervous system of c. elegans, Elife, 4 (2015), e12432. https://doi.org/10.7554/eLife.12432 doi: 10.7554/eLife.12432
|
| [41] |
D. L. Chase, M. R. Koelle, Biogenic amine neurotransmitters in c. elegans, WormBook: The Online Review of C. elegans Biology [Internet], 2007. https://doi.org/10.1895/wormbook.1.132.1 doi: 10.1895/wormbook.1.132.1
|
| [42] |
Z. F. Altun, B. Chen, Z.-W. Wang, D. H. Hall, High resolution map of caenorhabditis elegans gap junction proteins, Developmental dynamics: an official publication of the American Association of Anatomists, 238 (2009), 1936–1950. https://doi.org/10.1002/dvdy.22025 doi: 10.1002/dvdy.22025
|
| [43] | E. J. Jin, S. Park, X. Lyu, Y. Jin, Gap junctions: historical discoveries and new findings in the caenorhabditis elegans nervous system, Biol. Open, 9 (2020), bio053983. |
| [44] |
Q. Liu, G. Hollopeter, E. M. Jorgensen, Graded synaptic transmission at the caenorhabditis elegans neuromuscular junction, Proceedings of the National Academy of Sciences, 106 (2009), 10823–10828. https://doi.org/10.1073/pnas.0903570106 doi: 10.1073/pnas.0903570106
|
| [45] |
T. H. Lindsay, T. R. Thiele, S. R. Lockery, Optogenetic analysis of synaptic transmission in the central nervous system of the nematode caenorhabditis elegans, Nat. Commun., 2 (2011), 1–9. https://doi.org/10.1038/ncomms1304 doi: 10.1038/ncomms1304
|
| [46] |
A. Narayan, G. Laurent, P. W. Sternberg, Transfer characteristics of a thermosensory synapse in caenorhabditis elegans, Proceedings of the National Academy of Sciences, 108 (2011), 9667–9672. https://doi.org/10.1073/pnas.1106617108 doi: 10.1073/pnas.1106617108
|
| [47] |
P. Liu, B. Chen, Z.-W. Wang, Postsynaptic current bursts instruct action potential firing at a graded synapse, Nat. commun., 4 (2013), 1–12. https://doi.org/10.1038/ncomms2925 doi: 10.1038/ncomms2925
|
| [48] |
W. Zou, J. Fu, H. Zhang, K. Du, W. Huang, J. Yu, et al., Decoding the intensity of sensory input by two glutamate receptors in one c. elegans interneuron, Nat. commun., 9 (2018), 1–12. https://doi.org/10.1038/s41467-018-06819-5 doi: 10.1038/s41467-018-06819-5
|
| [49] |
R. E. Davis, A. O. Stretton, Passive membrane properties of motorneurons and their role in long-distance signaling in the nematode ascaris, J. Neurosci., 9 (1989), 403–414. https://doi.org/10.1523/JNEUROSCI.09-02-00403.1989 doi: 10.1523/JNEUROSCI.09-02-00403.1989
|
| [50] |
R. E. Davis, A. O. Stretton, Signaling properties of ascaris motorneurons: graded active responses, graded synaptic transmission, and tonic transmitter release, J. Neurosci., 9 (1989), 415–425. https://doi.org/10.1523/JNEUROSCI.09-02-00415.1989 doi: 10.1523/JNEUROSCI.09-02-00415.1989
|
| [51] |
S. R. Wicks, C. J. Roehrig, C. H. Rankin, A dynamic network simulation of the nematode tap withdrawal circuit: predictions concerning synaptic function using behavioral criteria, J. Neurosci., 16 (1996), 4017–4031. https://doi.org/10.1523/JNEUROSCI.16-12-04017.1996 doi: 10.1523/JNEUROSCI.16-12-04017.1996
|
| [52] |
P. Liu, B. Chen, R. Mailler, Z.-W. Wang, Antidromic-rectifying gap junctions amplify chemical transmission at functionally mixed electrical-chemical synapses, Nat. commun., 8 (2017), 14818. https://doi.org/10.1038/ncomms14818 doi: 10.1038/ncomms14818
|
| [53] |
Y. Shui, P. Liu, H. Zhan, B. Chen, Z.-W. Wang, Molecular basis of junctional current rectification at an electrical synapse, Sci. Adv., 6 (2020), eabb3076. https://doi.org/10.1126/sciadv.abb3076 doi: 10.1126/sciadv.abb3076
|
| [54] |
J. E. Mellem, P. J. Brockie, D. M. Madsen, A. V. Maricq, Action potentials contribute to neuronal signaling in c. elegans, Nat. neurosci., 11 (2008), 865–867. https://doi.org/10.1038/nn.2131 doi: 10.1038/nn.2131
|
| [55] |
D. Ramot, B. L. MacInnis, M. B. Goodman, Bidirectional temperature-sensing by a single thermosensory neuron in c. elegans, Nat. neurosci., 11 (2008), 908–915. https://doi.org/10.1038/nn.2157 doi: 10.1038/nn.2157
|
| [56] |
S. L. Geffeney, J. G. Cueva, D. A. Glauser, J. C. Doll, T. H. C. Lee, M. Montoya, et al., Deg/enac but not trp channels are the major mechanoelectrical transduction channels in a c. elegans nociceptor, Neuron, 71 (2011), 845–857. https://doi.org/10.1016/j.neuron.2011.06.038 doi: 10.1016/j.neuron.2011.06.038
|
| [57] |
M. Dobosiewicz, Q. Liu, C. I. Bargmann, Reliability of an interneuron response depends on an integrated sensory state, Elife, 8 (2019), e50566. https://doi.org/10.7554/eLife.50566 doi: 10.7554/eLife.50566
|
| [58] |
G. Schilardi, S. Kleinlogel, Two functional classes of rod bipolar cells in the healthy and degenerated optogenetically treated murine retina, Front. Cell. Neurosci., 15 (2021). https://doi.org/10.3389/fncel.2021.809531 doi: 10.3389/fncel.2021.809531
|
| [59] |
T. Aoyama, Y. Kamiyama, S. Usui, R. Blanco, C. F. Vaquero, P. de la Villa, Ionic current model of rabbit retinal horizontal cell, Neurosci. Res., 37 (2000), 141–151. https://doi.org/10.1016/S0168-0102(00)00111-5 doi: 10.1016/S0168-0102(00)00111-5
|
| [60] |
A. A. Koulakov, S. Raghavachari, A. Kepecs, J. E. Lisman, Model for a robust neural integrator, Nat. neurosci., 5 (2002), 775–782. https://doi.org/10.1038/nn893 doi: 10.1038/nn893
|
| [61] |
M. S. Goldman, J. H. Levine, G. Major, D. W. Tank, H. S. Seung, Robust persistent neural activity in a model integrator with multiple hysteretic dendrites per neuron, Cereb. cortex, 13 (2003), 1185–1195. https://doi.org/10.1093/cercor/bhg095 doi: 10.1093/cercor/bhg095
|
| [62] | M. Camperi, X. J. Wang, A model of visuospatial working memory in prefrontal cortex: recurrent network and cellular bistability, J. comput. neurosci., 5 (1998), 383–405. |
| [63] |
L. R. Varshney, B. L. Chen, E. Paniagua, D. H. Hall, D. B. Chklovskii, Structural properties of the caenorhabditis elegans neuronal network, PLoS Comput. Biol., 7 (2011), e1001066. https://doi.org/10.1371/journal.pcbi.1001066 doi: 10.1371/journal.pcbi.1001066
|
Figures(10) / Tables(2)

Loïs Naudin. Biological emergent properties in non-spiking neural networks[J]. AIMS Mathematics, 2022, 7(10): 19415-19439. doi: 10.3934/math.20221066







 DownLoad:
DownLoad: