Citation: Tingting Zhao, Robert J. Smith?. Global dynamical analysis of plant-disease models with nonlinear impulsive cultural control strategy[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 7022-7056. doi: 10.3934/mbe.2019353

| [1] | R. W. Gibson, J. P. Legg and G. W. Otim-Nape, Unusually severe symptoms are a characteristic of the current epidemic of mosaic virus disease of cassava in Uganda, Ann. Appl. Biol., 128 (1996), 479–490. |
| [2] | T. Iljon, J. Stirling and R. J. Smith?, A mathematical model describing an outbreak of Fire Blight, in Understanding the dynamics of emerging and re-emerging infectious diseases using mathematical models (eds. S. Mushayabasa and C.P. Bhunu), Transworld Research network, (2012), 91–104. |
| [3] | R. A. C. Jones, Using epidemiological information to develop effective integrated virus disease management strategies, Virus Res., 100 (2004), 5–30. |
| [4] | C. A. Gilligan, Sustainable agriculture and plant diseases: an epidemiological perspective, Phil. Trans. R. Soc. B, 363 (2008), 741–759. |
| [5] | L. M. C. Medina, I. T. Pacheco, R. G. G. Gonzalez, et al., Mathematical modeling tendencies in plant pathology, Afr. J. Biotechnol., 8 (2009), 7399–7408. |
| [6] | R. A. C. Jones, Determining threshold levels for seed-borne virus infection in seed stocks, Virus Res., 71 (2000), 171–183. |
| [7] | T. T. Zhao and S. Y. Tang, Plant disease control with Economic Threshold, J. Bioma., 24 (2009), 385–396. |
| [8] | F. Van den Bosch and A. M. Roos, The dynamics of infectious diseases in orchards with roguing and replanting as control strategy, J. Math. Biol., 35 (1996), 129–157. |
| [9] | M. S. Chan and M. J. Jeger, An analytical model of plant virus disease dynamics with roguing, J. Appl. Ecol., 31 (1994), 413–427. |
| [10] | H. R. Thieme and J. A. P. Heesterbeek, How to estimate the efficacy of periodic control of an infectious plant disease, Math. Biosci. 93 (1989), 15–29. |
| [11] | R. W. Gibson and V. Aritua, The perspective of sweet potato chlorotic stunt virus in sweet potato production in Africa, a review, Afr. Crop Sci. J., 10 (2002), 281–310. |
| [12] | J. M. Thresh and R. J. Cooter, Strategies for controlling cassava mosaic disease in Africa, Plant Pathol., 54 (2005), 587–614. |
| [13] | J. M. Thresh, The origins and epidemiology of some important plant virus diseases, Appl. Biol., 5 (1980), 1–65. |
| [14] | S. Fishman, R. Marcus, H. Talpaz, et al., Epidemiological and economic models for the spread and control of citrus tristeza virus disease, Phytoparasitica, 11 (1983), 39–49. |
| [15] | J. M. Thresh and G. K. Owusu, The control of cocoa swollen shoot disease in Ghana: an evaluation of eradication procedures, Crop Prot., 5 (1986) 41–52. |
| [16] | A. N. Adams, The incidence of plume pox virus in England and its control in orchards, in Plant Disease Epidemiology (ed. P.R. Scott and A. Bainbridge), Blackwell Scientific Publications, (1978), 213–219. |
| [17] | G. Hughes, N. McRoberts, L. V. Madden, et al., Validating mathematical models of plant-disease progress in space and time, Math. Med. Biol., 14 (1997), 85–112. |
| [18] | J. Holt and T. C. B. Chancellor, A model of plant virus disease epidemics in asynchronously-planted cropping systems, Plant Pathol., 46 (1997), 490–501. |
| [19] | F. van den Bosch, N. McRoberts, F. van den Berg, et al., The basic reproduction number of plant pathogens: matrix approaches to complex dynamics, Phytopathology, 98 (2008), 239–249. |
| [20] | C. Chen, Y. Kang and R. J. Smith?, Sliding motion and global dynamics of a Filippov fire-blight model with economic thresholds, Nonlinear Anal.-Real, 39 (2018), 492–519. |
| [21] | A. Wang, Y. Xiao and R. J. Smith?, Using non-smooth models to determine thresholds for microbial pest management, J. Math. Biol., 78 (2019), 1389–1424. |
| [22] | L. V. Madden, G. Hughes and F. van den Bosch, The study of plant disease epidemics, The American Phytopathological Society, St. Paul, 2007. |
| [23] | F. Van den Bosch, M. J. Jeger and C. A. Gilligan, Disease control and its selection for damaging plant virus strains in vegetatively propagated staple food crops; a theoretical assessment, Proc. R. Soc. B, 274 (2007), 11–18. |
| [24] | S. Fishman and R. Marcus, A model for spread of plant disease with periodic removals, J. Math. Biol., 21 (1984), 149–158. |
| [25] | Y. N. Xiao, D. Z. Cheng and H. S. Qin, Optimal impulsive control in periodic ecosystem, Syst. Contr. Lett., 55 (2006), 558–565. |
| [26] | X. Y. Zhang, Z. S. Shuai and K. Wang, Optimal impulsive harvesting policy for single population, Nonlinear Anal.-Real, 4 (2003), 639–651. |
| [27] | S. Y. Tang, Y. N. Xiao and R. A. Cheke, Dynamical analysis of plant disease models with cultural control strategies and economic thresholds, Math. Comput. Simul., 80 (2010), 894–921. |
| [28] | M. C. Smith, J. Holt, L. Kenyon, et al., Quantitative epidemiology of banana bunchy top virus disease and its control, Plant Pathol., 47 (1998), 177–187. |
| [29] | S. Y. Tang, Y. N. Xiao and R. A. Cheke, Multiple attractors of host-parasitoid models with integrated pest management strategies: eradication, persistence and outbreak, Theor. Popul. Biol., 73 (2008), 181–197. |
| [30] | T. T. Zhao, Y. N. Xiao and R. J. Smith?, Non-smooth plant disease models with economic thresholds, Math. Biosci., 241 (2013), 34–48. |
| [31] | S. Y. Tang, R. A. Cheke and Y. N. Xiao, Optimal impulsive harvesting on non-autonomous Beverton–Holt difference equations, Nonlinear Anal.-Theor., 65 (2006), 2311–2341. |
| [32] | D. Bainov and P. Simeonov, Impulsive Differential Equations: Periodic Solutions and Applications, Longman Scientific and Technical, New York, 1993. |
| [33] | J. M. Heffernan, R. J. Smith and L. M. Wahl, Perspectives on the basic reproductive ratio, J. R. Soc. Interface, 2 (2005), 281–293. |
| [34] | J. Li, D. Blakeley and R. J. Smith?, The Failure of R0, Comp. Math. Meth. Med., 2011 (2011), Article ID 527610. |
| [35] | A. Lakmech and O. Arino, Bifurcation of nontrival periodic solutions of impulsive differential equations arising chemotherapeutic treatment, Dyn. Cont. Discr. Impul. Syst., 7 (2000), 265–287. |
| [36] | M. D. McKay, W. J. Conover and R. J. Beckman, A comparison of three models for selecting values of input variables in the analysis of output from a computer code, Technometrics, 21 (1979), 239–245 . |
| [37] | R. M. Corless, G. H. Gonnet, D. E. G. Hare, et al., On the Lambert W Function, Adv. Comput. Math., 5 (1996), 329–359. |
| [38] | S. Y. Tang and Y. N. Xiao, Dynamics System of Single Species, Science Press, Beijing, 2008. |
| [39] | Y. A. Kuznetsov, Elements of Applied Bifurcation Theory, Springer-Verlag, New York, 2004. |
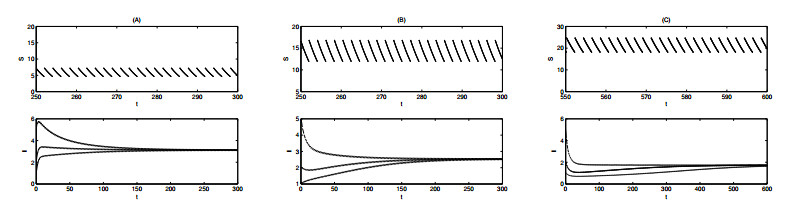
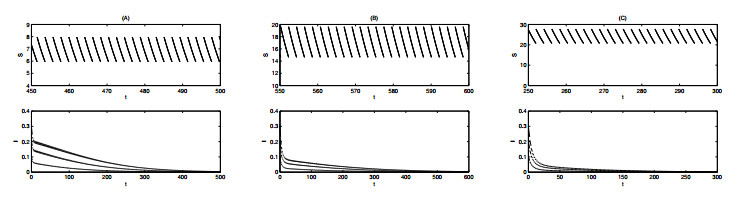
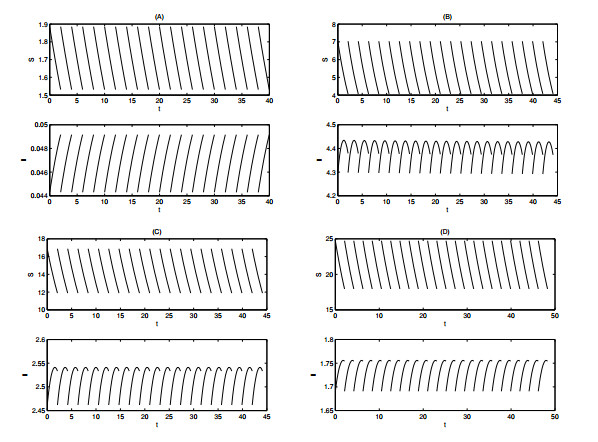
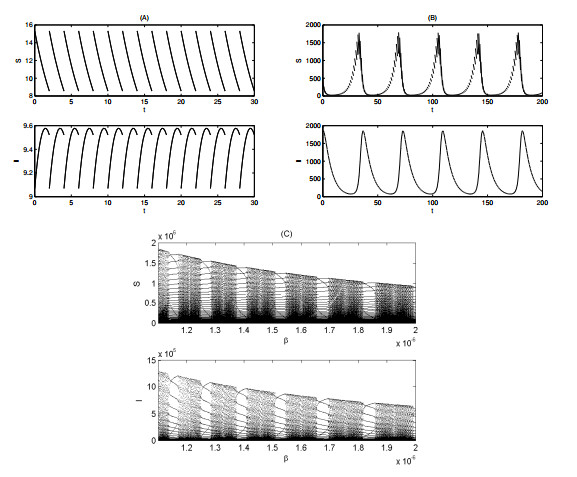
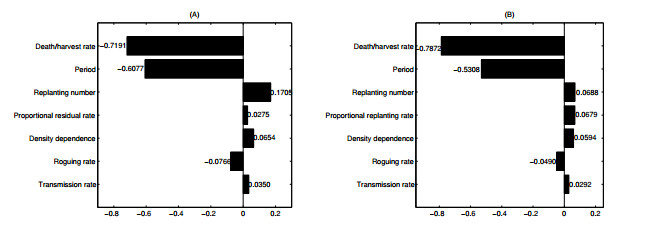
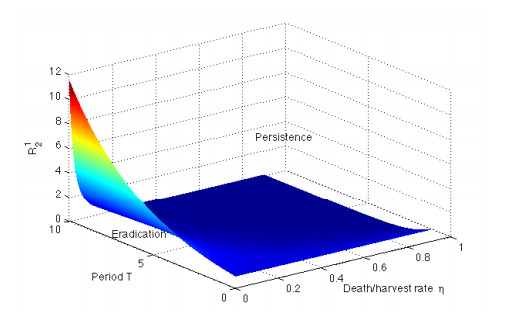
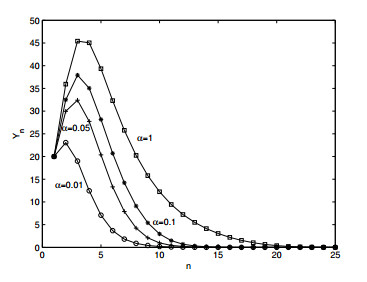
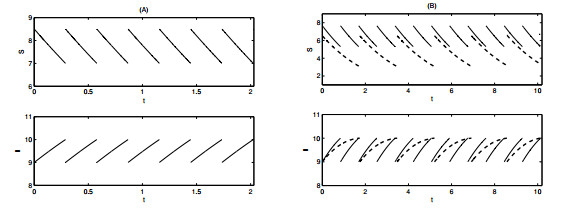
Figures(12) / Tables(1)

Tingting Zhao, Robert J. Smith?. Global dynamical analysis of plant-disease models with nonlinear impulsive cultural control strategy[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 7022-7056. doi: 10.3934/mbe.2019353







 DownLoad:
DownLoad: