Citation: Eman Koshlaf, Andrew S Ball. Soil bioremediation approaches for petroleum hydrocarbon polluted environments[J]. AIMS Microbiology, 2017, 3(1): 25-49. doi: 10.3934/microbiol.2017.1.25

| [1] | Petroleum B, BP statistical review of world energy, 2015. Available from: www. bp. com/statisticalreview. |
| [2] | Sihag S, Pathak H, Jaroli D (2014) Factors affecting the rate of biodegradation of polyaromatic hydrocarbons. Int J Pure App Biosci 2: 185-202. |
| [3] |
Stroud JL, Paton GI, Semple KT (2007) Microbe-aliphatic hydrocarbon interactions in soil: implications for biodegradation and bioremediation. J Appl Microbiol 102: 1239-1253. doi: 10.1111/j.1365-2672.2007.03401.x

|
| [4] | Das N, Chandran P (2011) Microbial degradation of petroleum hydrocarbon contaminants: an overview. Biotechnol Res Int 2011: 941810. |
| [5] |
Gallego JL, Loredo J, Llamas JF, et al. (2001) Bioremediation of diesel-contaminated soils: evaluation of potential in situ techniques by study of bacterial degradation. Biodegradation 12: 325-335. doi: 10.1023/A:1014397732435
|
| [6] | Gong R, Hochmuth M, Weatherburn DC (2003) Polycyclic aromatic hydrocarbons and diesel particulates. |
| [7] |
Latif I, Karim A, Zuki A, et al. (2010) Pulmonary modulation of benzo [a] pyrene-induced hemato-and hepatotoxicity in broilers. Poultry Sci 89: 1379-1388. doi: 10.3382/ps.2009-00622
|
| [8] |
Samanta SK, Singh OV, Jain RK (2002) Polycyclic aromatic hydrocarbons: environmental pollution and bioremediation. Trends Biotechnol 20: 243-248. doi: 10.1016/S0167-7799(02)01943-1
|
| [9] |
Huang XD, El-Alawi Y, Penrose DM, et al. (2004) A multi-process phytoremediation system for removal of polycyclic aromatic hydrocarbons from contaminated soils. Environ Pollut 130: 465-476. doi: 10.1016/j.envpol.2003.09.031
|
| [10] |
Arthur EL, Rice PJ, Rice PJ, et al. (2005) Phytoremediation—An Overview. CRC Crit Rev Plant Sci 24: 109-122. doi: 10.1080/07352680590952496
|
| [11] |
Guo J, Feng R, Ding Y, et al. (2014) Applying carbon dioxide, plant growth-promoting rhizobacterium and EDTA can enhance the phytoremediation efficiency of ryegrass in a soil polluted with zinc, arsenic, cadmium and lead. J Environ Manage 141: 1-8. doi: 10.1016/j.jenvman.2013.12.039
|
| [12] | Onwurah I, Ogugua V, Onyike N, et al. (2007) Crude oil spills in the environment, effects and some innovative clean-up biotechnologies. |
| [13] |
Yemashova NA, Murygina VP, Zhukov DV, et al. (2007) Biodeterioration of crude oil and oil derived products: a review. Rev Environ Sci Biotechnol 6: 315-337. doi: 10.1007/s11157-006-9118-8
|
| [14] |
Head IM, Jones DM, Röling WF (2006) Marine microorganisms make a meal of oil. Nat Rev Microbiol 4: 173-182. doi: 10.1038/nrmicro1348
|
| [15] | Yasin G, Bhanger MI, Ansari TM, et al. (2013) Quality and chemistry of crude oils. J Pet Technol Altern Fuels 4: 53-63. |
| [16] | Alvarez PJ, Illman WA (2005) Bioremediation and natural attenuation: process fundamentals and mathematical models. John Wiley & Sons. |
| [17] |
Martinez-Gomez C, Vethaak A, Hylland K, et al. (2010) A guide to toxicity assessment and monitoring effects at lower levels of biological organization following marine oil spills in European waters. ICES J Mar Sci 67: 1105-1118. doi: 10.1093/icesjms/fsq017
|
| [18] |
Haritash A, Kaushik C (2009) Biodegradation aspects of polycyclic aromatic hydrocarbons (PAHs): a review. J Hazard Mater 169: 1-15. doi: 10.1016/j.jhazmat.2009.03.137
|
| [19] |
Mishra S, Jyot J, Kuhad RC, et al. (2001) Evaluation of inoculum addition to stimulate in situ bioremediation of oily-sludge-contaminated soil. Appl Environ Microb 67: 1675-1681. doi: 10.1128/AEM.67.4.1675-1681.2001
|
| [20] |
Kim KH, Jahan SA, Kabir E, et al. (2013) A review of airborne polycyclic aromatic hydrocarbons (PAHs) and their human health effects. Environ Int 60: 71-80. doi: 10.1016/j.envint.2013.07.019
|
| [21] | Masih J, Singhvi R, Kumar K, et al. (2012) Seasonal variation and sources of polycyclic aromatic hydrocarbons (PAHs) in indoor and outdoor air in a semi arid tract of northern India. AAQR 12: 515-525. |
| [22] | Perelo LW (2010) Review: in situ and bioremediation of organic pollutants in aquatic sediments. J Hazard Mater 177: 81-89. |
| [23] |
Gaspar A, Zellermann E, Lababidi S, et al. (2012) Characterization of saturates, aromatics, resins, and asphaltenes heavy crude oil fractions by atmospheric pressure laser ionization Fourier transform ion cyclotron resonance mass spectrometry. Energ Fuel 26: 3481-3487. doi: 10.1021/ef3001407
|
| [24] |
Leyva C, Ancheyta J, Berrueco C, et al. (2013) Chemical characterization of asphaltenes from various crude oils. Fuel Process Technol 106: 734-738. doi: 10.1016/j.fuproc.2012.10.009
|
| [25] |
Vinas M, Grifoll M, Sabate J, et al. (2002) Biodegradation of a crude oil by three microbial consortia of different origins and metabolic capabilities. J Ind Microbiol Biot 28: 252-260. doi: 10.1038/sj.jim.7000236
|
| [26] |
He L, Lin F, Li X, et al. (2015) Interfacial sciences in unconventional petroleum production: from fundamentals to applications. Chem Soc Rev 44: 5446-5494. doi: 10.1039/C5CS00102A
|
| [27] |
Fuentes S, Méndez V, Aguila P, et al. (2014) Bioremediation of petroleum hydrocarbons: catabolic genes, microbial communities, and applications. Appl Microbiol Biot 98: 4781-4794. doi: 10.1007/s00253-014-5684-9
|
| [28] |
Dong TT, Lee BK (2009) Characteristics, toxicity, and source apportionment of polycylic aromatic hydrocarbons (PAHs) in road dust of Ulsan, Korea. Chemosphere 74: 1245-1253. doi: 10.1016/j.chemosphere.2008.11.035
|
| [29] |
Ou S, Zheng J, Zheng J, et al. (2004) Petroleum hydrocarbons and polycyclic aromatic hydrocarbons in the surficial sediments of Xiamen Harbour and Yuan Dan Lake, China. Chemosphere 56: 107-112. doi: 10.1016/j.chemosphere.2004.02.022
|
| [30] |
Chandra S, Sharma R, Singh K, et al. (2013) Application of bioremediation technology in the environment contaminated with petroleum hydrocarbon. Ann Microbiol 63: 417-431. doi: 10.1007/s13213-012-0543-3
|
| [31] |
Paruk JD, Adams EM, Uher-Koch H, et al. (2016) Polycyclic aromatic hydrocarbons in blood related to lower body mass in common loons. Sci Total Environ 565: 360-368. doi: 10.1016/j.scitotenv.2016.04.150
|
| [32] |
Militon C, Boucher D, Vachelard C, et al. (2010) Bacterial community changes during bioremediation of aliphatic hydrocarbon-contaminated soil. Fems Microbiol Ecol 74: 669-681. doi: 10.1111/j.1574-6941.2010.00982.x
|
| [33] |
Wasmund K, Burns KA, Kurtböke DI, et al. (2009) Novel alkane hydroxylase gene (alkB) diversity in sediments associated with hydrocarbon seeps in the Timor Sea, Australia. Appl Environ Microb 75: 7391-7398. doi: 10.1128/AEM.01370-09
|
| [34] |
Adgate JL, Goldstein BD, McKenzie LM (2014) Potential public health hazards, exposures and health effects from unconventional natural gas development. Environ Sci Technol 48: 8307-8320. doi: 10.1021/es404621d
|
| [35] | Zhu X, Venosa AD, Suidan MT, et al. (2001) Guidelines for the bioremediation of marine shorelines and freshwater wetlands. US EPA |
| [36] | Sadler R, Connell D (2003) Analytical methods for the determination of total petroleum hydrocarbons in soil. In: Proceedings of the fifth national workshop on the assessment of site contamination, 133-150. |
| [37] | Van Beilen JB, Funhoff EG (2007) Alkane hydroxylases involved in microbial alkane degradation. Appl Microbiol Biot 74: 13-21. |
| [38] |
Hatzinger PB, Alexander M (1995) Effect of aging of chemicals in soil on their biodegradability and extractability. Environ Sci Technol 29: 537-545. doi: 10.1021/es00002a033
|
| [39] |
Semple KT, Morriss A, Paton G (2003) Bioavailability of hydrophobic organic contaminants in soils: fundamental concepts and techniques for analysis. Eur J Soil Sci 54: 809-818. doi: 10.1046/j.1351-0754.2003.0564.x
|
| [40] |
Baboshin M, Golovleva L (2012) Aerobic bacterial degradation of polycyclic aromatic hydrocarbons (PAHs) and its kinetic aspects. Microbiology 81: 639-650. doi: 10.1134/S0026261712060021
|
| [41] |
Rojo F (2009) Degradation of alkanes by bacteria. Environ Microbiol 11: 2477-2490. doi: 10.1111/j.1462-2920.2009.01948.x
|
| [42] | Wentzel A, Ellingsen TE, Kotlar HK, et al. (2007) Bacterial metabolism of long-chain n-alkanes. Appl Microbiol Biot 76: 1209-1221. |
| [43] | Kotani T, Yurimoto H, Kato N, et al. (2007) Novel acetone metabolism in a propane-utilizing bacterium, Gordonia sp. strain TY-5. J Bacteriol 189: 886-893. |
| [44] |
Bamforth SM, Singleton I (2005) Bioremediation of polycyclic aromatic hydrocarbons: current knowledge and future directions. J Chem Technol Biot 80: 723-736. doi: 10.1002/jctb.1276
|
| [45] |
Kanaly RA, Harayama S (2010) Advances in the field of high-molecular-weight polycyclic aromatic hydrocarbon biodegradation by bacteria. Microb Biotechnol 3: 136-164. doi: 10.1111/j.1751-7915.2009.00130.x
|
| [46] |
Seo JS, Keum YS, Li QX (2009) Bacterial degradation of aromatic compounds. Int J Environ Res Public Health 6: 278-309. doi: 10.3390/ijerph6010278
|
| [47] | Maigari AU, Maigari MU (2015) Microbial metabolism of polycyclic aromatic hydrocarbons (PAHs): a review. Int J Sci Eng 6: 1449-1459. |
| [48] | Rojo F, (2010) Enzymes for aerobic degradation of alkanes, In: Handbook of hydrocarbon and lipid microbiology, Springer, 781-797. |
| [49] |
Tortella GR, Diez MC, Durán N (2005) Fungal diversity and use in decomposition of environmental pollutants. Crit Rev Microbiol 31: 197-212. doi: 10.1080/10408410500304066
|
| [50] |
Brooijmans RJ, Pastink MI, Siezen RJ (2009) Hydrocarbon-degrading bacteria: the oil-spill clean-up crew. Microb Biotechnol 2: 587-594. doi: 10.1111/j.1751-7915.2009.00151.x
|
| [51] |
Germida J, Frick C, Farrell R, et al. (2002) Phytoremediation of oil-contaminated soils. Dev Soil Sci 28: 169-186. doi: 10.1016/S0166-2481(02)80015-0
|
| [52] | Wang YN, Cai H, Chi CQ, et al. (2007) Halomonas shengliensis sp. nov., a moderately halophilic, denitrifying, crude-oil-utilizing bacterium. Int J Syst Evol Micr 57: 1222-1226. |
| [53] | Mnif S, Chamkha M, Sayadi S (2009) Isolation and characterization of Halomonas sp. strain C2SS100, a hydrocarbon-degrading bacterium under hypersaline conditions. J Appl Microbiol 107: 785-794. |
| [54] |
Borzenkov I, Milekhina E, Gotoeva M, et al. (2006) The properties of hydrocarbon-oxidizing bacteria isolated from the oilfields of Tatarstan, Western Siberia, and Vietnam. Microbiology 75: 66-72. doi: 10.1134/S0026261706010127
|
| [55] | Dastgheib SMM, Amoozegar MA, Khajeh K, et al. (2011) A halotolerant Alcanivorax sp. strain with potential application in saline soil remediation. Appl Microbiol Biot 90: 305-312. |
| [56] |
Chaillan F, Le Flèche A, Bury E, et al. (2004) Identification and biodegradation potential of tropical aerobic hydrocarbon-degrading microorganisms. Res Microbiol 155: 587-595. doi: 10.1016/j.resmic.2004.04.006
|
| [57] |
Moody JD, Freeman JP, Fu PP, et al. (2004) Degradation of benzo [a] pyrene by Mycobacterium vanbaalenii PYR-1. Appl Environ Microb 70: 340-345. doi: 10.1128/AEM.70.1.340-345.2004
|
| [58] |
Van Beilen JB, Li Z, Duetz WA, et al. (2003) Diversity of alkane hydroxylase systems in the environment. Oil Gas Sci Technol 58: 427-440. doi: 10.2516/ogst:2003026
|
| [59] |
McKew BA, Coulon F, Osborn AM, et al. (2007) Determining the identity and roles of oil-metabolizing marine bacteria from the Thames estuary, UK. Environ Microbiol 9: 165-176. doi: 10.1111/j.1462-2920.2006.01125.x
|
| [60] |
Pilon-Smits E (2005) Phytoremediation. Annu Rev Plant Biol 56: 15-39. doi: 10.1146/annurev.arplant.56.032604.144214
|
| [61] |
Holden P, LaMontagne M, Bruce A, et al. (2002) Assessing the role of Pseudomonas aeruginosa surface-active gene expression in hexadecane biodegradation in sand. Appl Environ Microb 68: 2509-2518. doi: 10.1128/AEM.68.5.2509-2518.2002
|
| [62] |
Makadia TH, Adetutu EM, Simons KL, et al. (2011) Re-use of remediated soils for the bioremediation of waste oil sludge. J Environ Manage 92: 866-871. doi: 10.1016/j.jenvman.2010.10.059
|
| [63] |
Sheppard PJ, Simons KL, Kadali KK, et al. (2012) The importance of weathered crude oil as a source of hydrocarbonoclastic microorganisms in contaminated seawater. J Microbiol Biotechnol 22: 1185-1192. doi: 10.4014/jmb.1201.01049
|
| [64] | Rodríguez-Blanco A, Antoine V, Pelletier E, et al. (2010) Effects of temperature and fertilization on total vs. active bacterial communities exposed to crude and diesel oil pollution in NW Mediterranean Sea. Environ Pollut 158: 663-673. |
| [65] |
Sarkar D, Ferguson M, Datta R, et al. (2005) Bioremediation of petroleum hydrocarbons in contaminated soils: comparison of biosolids addition, carbon supplementation, and monitored natural attenuation. Environ Pollut 136: 187-195. doi: 10.1016/j.envpol.2004.09.025
|
| [66] |
Tang J, Wang R, Niu X, et al. (2010) Enhancement of soil petroleum remediation by using a combination of ryegrass (Lolium perenne) and different microorganisms. Soil Till Res 110: 87-93. doi: 10.1016/j.still.2010.06.010
|
| [67] |
Li X, Wu Y, Lin X, et al. (2012) Dissipation of polycyclic aromatic hydrocarbons (PAHs) in soil microcosms amended with mushroom cultivation substrate. Soil Biol Biochem 47: 191-197. doi: 10.1016/j.soilbio.2012.01.001
|
| [68] | Agnello AC, Bagard M, Van Hullebusch ED, et al. (2016) Comparative bioremediation of heavy metals and petroleum hydrocarbons co-contaminated soil by natural attenuation, phytoremediation, bioaugmentation and bioaugmentation-assisted phytoremediation. Sci Total Environ 563: 693-703. |
| [69] |
Kadali KK, Simons KL, Sheppard PJ, et al. (2012) Mineralisation of weathered crude oil by a hydrocarbonoclastic consortia in marine mesocosms. Water Air Soil Poll 223: 4283-4295. doi: 10.1007/s11270-012-1191-8
|
| [70] |
Yu K, Wong A, Yau K, et al. (2005) Natural attenuation, biostimulation and bioaugmentation on biodegradation of polycyclic aromatic hydrocarbons (PAHs) in mangrove sediments. Mar Pollut Bull 51: 1071-1077. doi: 10.1016/j.marpolbul.2005.06.006
|
| [71] |
Margesin R, Hämmerle M, Tscherko D (2007) Microbial activity and community composition during bioremediation of diesel-oil-contaminated soil: effects of hydrocarbon concentration, fertilizers, and incubation time. Microbial Ecol 53: 259-269. doi: 10.1007/s00248-006-9136-7
|
| [72] |
Festa S, Coppotelli B, Morelli I (2016) Comparative bioaugmentation with a consortium and a single strain in a phenanthrene-contaminated soil: Impact on the bacterial community and biodegradation. Appl Soil Ecol 98: 8-19. doi: 10.1016/j.apsoil.2015.08.025
|
| [73] | Nikolopoulou M, Kalogerakis N, (2010) Biostimulation strategies for enhanced bioremediation of marine oil spills including chronic pollution. In: Handbook of hydrocarbon and lipid microbiology, Springer, 2521-2529. |
| [74] | Alexander M (1999) Biodegradation and bioremediation. Gulf Professional Publishing. |
| [75] |
Walworth J, Pond A, Snape I, et al. (2007) Nitrogen requirements for maximizing petroleum bioremediation in a sub-Antarctic soil. Cold Reg Sci Technol 48: 84-91. doi: 10.1016/j.coldregions.2006.07.001
|
| [76] |
Piehler MF, Swistak J, Pinckney J, et al. (1999) Stimulation of diesel fuel biodegradation by indigenous nitrogen fixing bacterial consortia. Microbial Ecol 38: 69-78. doi: 10.1007/s002489900157
|
| [77] |
Li H, Zhao Q, Boufadel MC, et al. (2007) A universal nutrient application strategy for the bioremediation of oil-polluted beaches. Mar Pollut Bull 54: 1146-1161. doi: 10.1016/j.marpolbul.2007.04.015
|
| [78] |
Chaillan F, Chaineau C, Point V, et al. (2006) Factors inhibiting bioremediation of soil contaminated with weathered oils and drill cuttings. Environ Pollut 144: 255-265. doi: 10.1016/j.envpol.2005.12.016
|
| [79] |
Sayara T, Borràs E, Caminal G, et al. (2011) Bioremediation of PAHs-contaminated soil through composting: Influence of bioaugmentation and biostimulation on contaminant biodegradation. Int Biodeter Biodegr 65: 859-865. doi: 10.1016/j.ibiod.2011.05.006
|
| [80] |
Ros M, Rodriguez I, Garcia C, et al. (2010) Microbial communities involved in the bioremediation of an aged recalcitrant hydrocarbon polluted soil by using organic amendments. Bioresour Technol 101: 6916-6923. doi: 10.1016/j.biortech.2010.03.126
|
| [81] |
Blyth W, Shahsavari E, Morrison PD, et al. (2015) Biosurfactant from red ash trees enhances the bioremediation of PAH contaminated soil at a former gasworks site. J Environ Manage 162: 30-36. doi: 10.1016/j.jenvman.2015.07.041
|
| [82] |
Mair J, Schinner F, Margesin R (2013) A feasibility study on the bioremediation of hydrocarbon-contaminated soil from an alpine former military site: effects of temperature and biostimulation. Cold Reg Sci Technol 96: 122-128. doi: 10.1016/j.coldregions.2013.07.006
|
| [83] |
Liu W, Luo Y, Teng Y, et al. (2010) Bioremediation of oily sludge-contaminated soil by stimulating indigenous microbes. Environ Gecochem Hlth 32: 23-29. doi: 10.1007/s10653-009-9262-5
|
| [84] |
Palmroth MR, Pichtel J, Puhakka JA (2002) Phytoremediation of subarctic soil contaminated with diesel fuel. Bioresour Technol 84: 221-228. doi: 10.1016/S0960-8524(02)00055-X
|
| [85] |
Schaefer M, Juliane F (2007) The influence of earthworms and organic additives on the biodegradation of oil contaminated soil. Appl Soil Ecol 36: 53-62. doi: 10.1016/j.apsoil.2006.11.002
|
| [86] |
Suja F, Rahim F, Taha MR, et al. (2014) Effects of local microbial bioaugmentation and biostimulation on the bioremediation of total petroleum hydrocarbons (TPH) in crude oil contaminated soil based on laboratory and field observations. Int Biodeter Biodegr 90: 115-122. doi: 10.1016/j.ibiod.2014.03.006
|
| [87] | Al-Sulaimani HS, Al-Wahaibi YM, Al-Bahry S, et al. (2010) Experimental investigation of biosurfactants produced by Bacillus species and their potential for MEOR in Omani oil field. Society of Petroleum Engineers. |
| [88] |
Reid BJ, Jones KC, Semple KT (2000) Bioavailability of persistent organic pollutants in soils and sediments—a perspective on mechanisms, consequences and assessment. Environ Pollut 108: 103-112. doi: 10.1016/S0269-7491(99)00206-7
|
| [89] |
Liu PWG, Chang TC, Chen CH, et al. (2014) Bioaugmentation efficiency investigation on soil organic matters and microbial community shift of diesel-contaminated soils. Int Biodeter Biodegr 95: 276-284. doi: 10.1016/j.ibiod.2014.05.004
|
| [90] |
Hyun S, Ahn MY, Zimmerman AR, et al. (2008) Implication of hydraulic properties of bioremediated diesel-contaminated soil. Chemosphere 71: 1646-1653. doi: 10.1016/j.chemosphere.2008.01.026
|
| [91] |
Nam K, Chung N, Alexander M (1998) Relationship between organic matter content of soil and the sequestration of phenanthrene. Environ Sci Technol 32: 3785-3788. doi: 10.1021/es980428m
|
| [92] | Alexander M (2000) Aging, bioavailability, and overestimation of risk from environmental pollutants. Environ Sci Technol 34: 4259-4265. |
| [93] |
Si-Zhong Y, Hui-Jun J, Zhi W, et al. (2009) Bioremediation of oil spills in cold environments: a review. Pedosphere 19: 371-381. doi: 10.1016/S1002-0160(09)60128-4
|
| [94] | Okoh AI (2006) Biodegradation alternative in the cleanup of petroleum hydrocarbon pollutants. Biotechnol Mol Biol Rev 1: 38-50. |
| [95] | Margesin R, Schinner F (2001) Biodegradation and bioremediation of hydrocarbons in extreme environments. Appl Microbiol Biot 56: 650-663. |
| [96] |
Perfumo A, Banat IM, Marchant R, et al. (2007) Thermally enhanced approaches for bioremediation of hydrocarbon-contaminated soils. Chemosphere 66: 179-184. doi: 10.1016/j.chemosphere.2006.05.006
|
| [97] |
Scherr K, Aichberger H, Braun R, et al. (2007) Influence of soil fractions on microbial degradation behavior of mineral hydrocarbons. Eur J Soil Biol 43: 341-350. doi: 10.1016/j.ejsobi.2007.03.009
|
| [98] |
Amellal N, Portal J-M, Berthelin J (2001) Effect of soil structure on the bioavailability of polycyclic aromatic hydrocarbons within aggregates of a contaminated soil. Appl Geochem 16: 1611-1619. doi: 10.1016/S0883-2927(01)00034-8
|
| [99] |
Chaerun S, Tazaki K (2005) How kaolinite plays an essential role in remediating oil-polluted seawater. Clay Miner 40: 481-491. doi: 10.1180/0009855054040185
|
| [100] | Atlas RM, Philp J (2005) Bioremediation. Applied microbial solutions for real-world environmental cleanup. ASM Press. |
| [101] |
Zucchi M, Angiolini L, Borin S, et al. (2003) Response of bacterial community during bioremediation of an oil-polluted soil. J Appl Microbiol 94: 248-257. doi: 10.1046/j.1365-2672.2003.01826.x
|
| [102] | Tang J, Wang M, Wang F, et al. (2011) Eco-toxicity of petroleum hydrocarbon contaminated soil. J Environ Sci 23: 845-851. |
| [103] |
Sheppard PJ, Adetutu EM, Makadia TH, et al. (2011) Microbial community and ecotoxicity analysis of bioremediated, weathered hydrocarbon-contaminated soil. Soil Research 49: 261-269. doi: 10.1071/SR10159
|
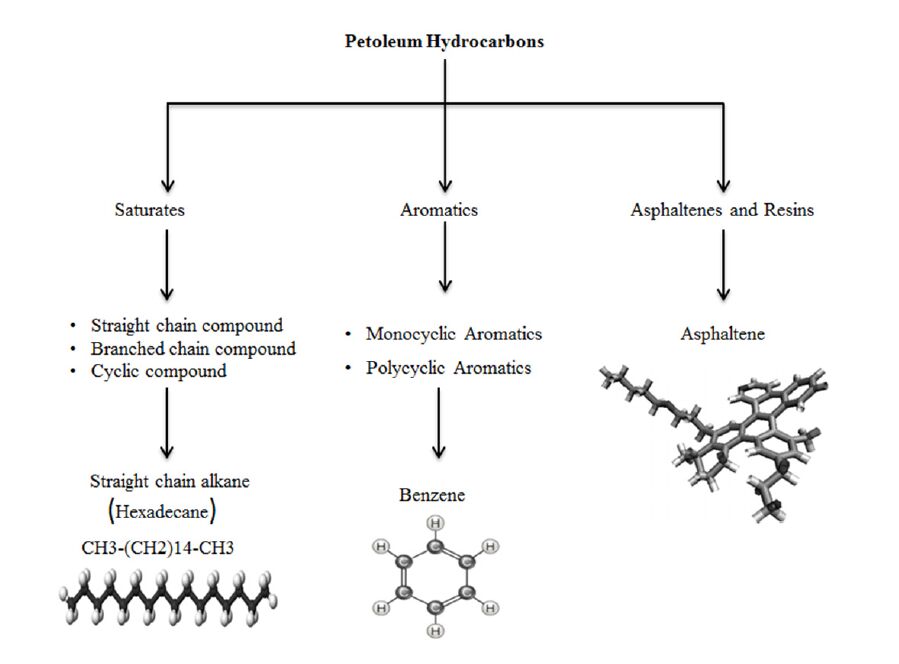

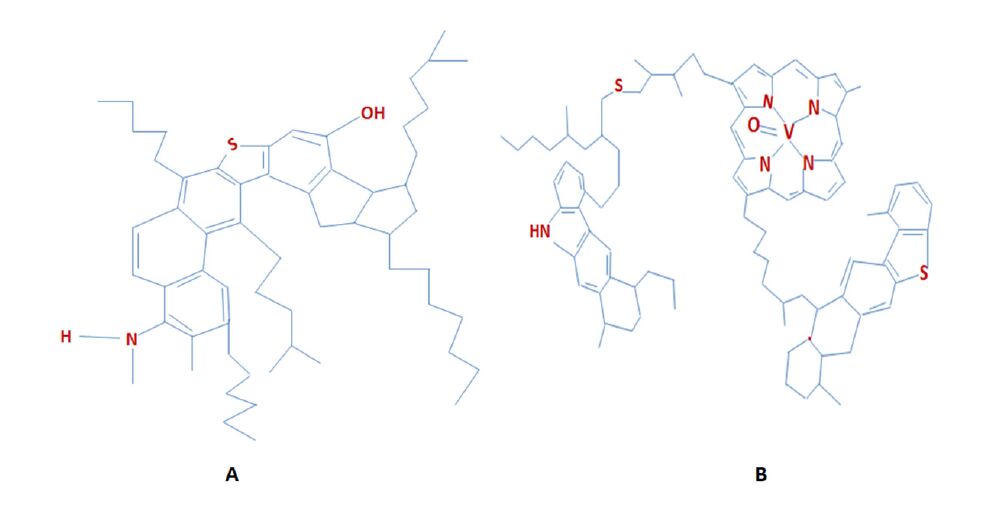
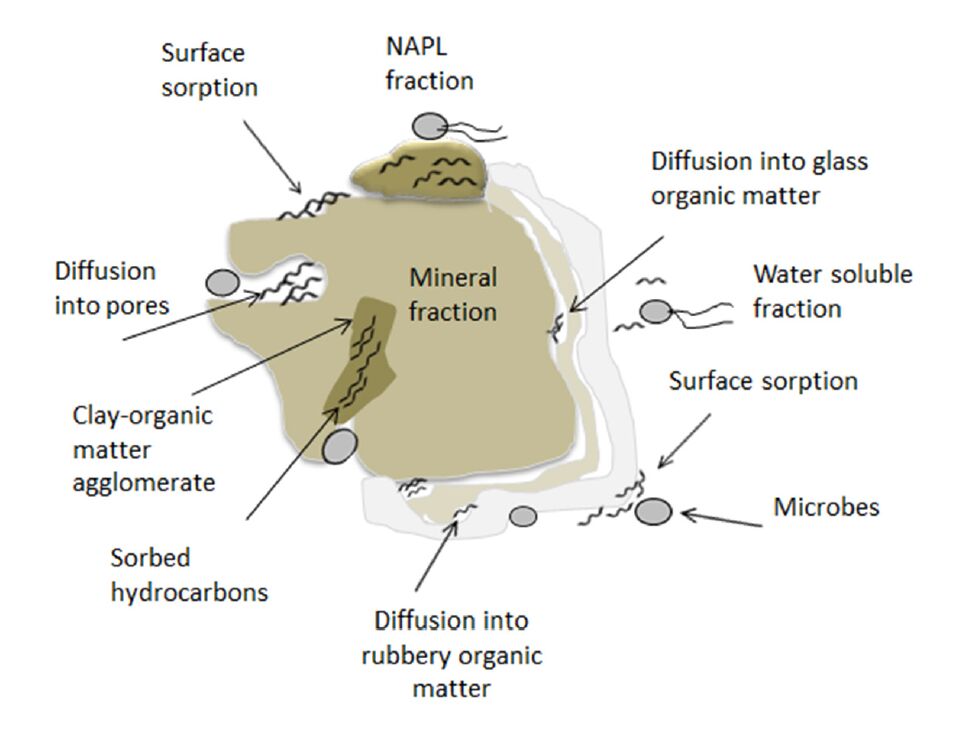
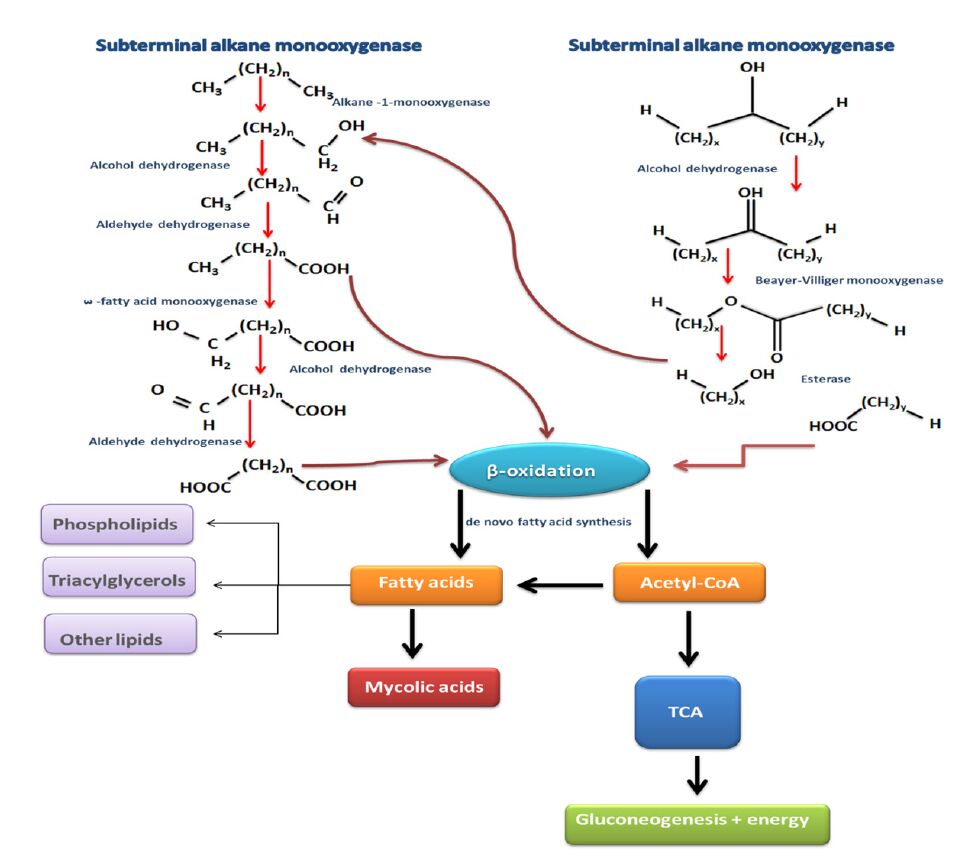

Figures(6) / Tables(4)

Eman Koshlaf, Andrew S Ball. Soil bioremediation approaches for petroleum hydrocarbon polluted environments[J]. AIMS Microbiology, 2017, 3(1): 25-49. doi: 10.3934/microbiol.2017.1.25







 DownLoad:
DownLoad: