Citation: Ziyu Wu, Sugui Wang, Qiang Li, Qingsong Zhao, Mingming Shao. Identification of 10 differently expressed lncRNAs as prognostic biomarkers for prostate adenocarcinoma[J]. Mathematical Biosciences and Engineering, 2020, 17(3): 2037-2047. doi: 10.3934/mbe.2020108

| [1] | X. Z. Wang, J. R. Beebe, L. Pwiti, A. Bielawska, M. J. Smyth, Aberrant sphingolipid signaling is involved in the resistance of prostate cancer cell lines to chemotherapy, Cancer Res., 59 (1999), 5842-5848. |
| [2] | K. E. Richert-Boe, L. L. Humphrey, A. G. Glass, N. S. Weiss, Screening digital rectal examination and prostate cancer mortality: A case-control study, J. Med. Screening, 5 (1998), 99-103. |
| [3] | W. Zhou, Z. Tao, Z. Wang, W. Hu, M. Shen, L. Zhou, et al., Long noncoding RNA PCA3 gene promoter region is related to the risk of prostate cancer on Chinese males, Exp. Mol. Pathol., 97 (2014), 550-553. |
| [4] | C. Haldrup, A. L. Pedersen, N. Øgaard, S. H. Strand, S. Høyer, M. Borre, et al., Biomarker potential of ST6GALNAC3 and ZNF660 promoter hypermethylation in prostate cancer tissue and liquid biopsies, Mol. Oncol., 12 (2018), 545-560. |
| [5] | L. J. Fan, H. J. Han, J. Guan, X. W. Zhang, Q. H. Cui, H. Shen, et al., Aberrantly expressed long noncoding RNAs in recurrent implantation failure: A microarray related study, Syst. Biol. Reprod. Med., 63 (2017), 269-278. |
| [6] | L. Deng, S. B. Yang, F. F. Xu, J. H. Zhang, Long noncoding RNA CCAT1 promotes hepatocellular carcinoma progression by functioning as let-7 sponge, J. Exp. Clin. Cancer Res., 34 (2015), 18. |
| [7] | D. Chakravarty, A. Sboner, S. S. Nair, E. Giannopoulou, R. Li, S. Hennig, et al., The oestrogen receptor alpha-regulated lncRNA NEAT1 is a critical modulator of prostate cancer, Nat. Communi., 5 (2014), 5383. |
| [8] | W. C. Cui, Y. F. Wu, H. M. Qu, Up-regulation of long non-coding RNA PCAT-1 correlates with tumor progression and poor prognosis in gastric cancer, Eur. Rev. Med. Pharmacol. Sci., 21 (2017), 3021-3027. |
| [9] | W. H. Shi, Q. Q. Wu, S. Q. Li, T. X. Yang, Z. H. Liu, Y. S. Tong, et al., Upregulation of the long noncoding RNA PCAT-1 correlates with advanced clinical stage and poor prognosis in esophageal squamous carcinoma, Tumour Biol., 36 (2015), 2501-2507. |
| [10] | X. Wan, W. Huang, S. Yang, Y. Zhang, H. Pu, F. Fu, et al., Identification of androgen-responsive lncRNAs as diagnostic and prognostic markers for prostate cancer, Oncotarget, 7 (2016), 60503-60518. |
| [11] | K. Tomczak, P. Czerwinska, M. Wiznerowicz, The Cancer Genome Atlas (TCGA): An immeasurable source of knowledge, Contemp. Oncol., 19 (2015), A68-A77. |
| [12] | J. Du, Z. Yuan, Z. Ma, J. Song, X. Xie, Y. Chen, KEGG-PATH: Kyoto encyclopedia of genes and genomes-based pathway analysis using a path analysis model, Mol. Biosyst., 10 (2014), 2441-2447. |
| [13] | Z. Tang, C. Li, B. Kang, G. Gao, C. Li, Z. Zhang, GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses, Nucleic Acids Res., 45 (2017), W98-W102. |
| [14] | Z. Zhou, Z. Dai, S. Zhou, Z. Hu, Q. Chen, Y. Zhao, et al., HNRNPAB induces epithelial-mesenchymal transition and promotes metastasis of hepatocellular carcinoma by transcriptionally activating SNAIL, Cancer Res., 74 (2014), 2750-2762. |
| [15] | Y. Zhang, S. Pitchiaya, M. Cieślik, Y. S. Niknafs, J. Tien, Y. Hosono, et al., Analysis of the androgen receptor-regulated lncRNA landscape identifies a role for ARLNC1 in prostate cancer progression, Nat. Genet., 50 (2018), 814-824. |
| [16] | F. F. Coelho, F. L. Guimarã£Es, W. L. Cabral, P. G. Salles, E. C. Mateo, L. M. N. Nogueita, et al., Expression of PCA3 and PSA genes as a biomarker for differential diagnosis of nodular hyperplasia and prostate cancer, Genet. Mol. Res., 14 (2015), 13519-13531. |
| [17] | C. Huang, W. Yu, Q. Wang, H. Cui, Y. Wang, L. Zhang, et al., Increased expression of the lncRNA PVT1 is associated with poor prognosis in pancreatic cancer patients, Minerva Med., 106 (2015), 143-149. |
| [18] | M. Wu, X. Li, C. K. Kwoh, S. K. Ng, A core-attachment based method to detect protein complexes in PPI networks, BMC Bioinf., 10 (2009), 169. |
| [19] | F. Lan, R. E. Colins, R. De Cegli, R. Alpatov, J. R. Horton, X. Shi, et al., Recognition of unmethylated histone H3 lysine 4 links BHC80 to LSD1-mediated gene repression, Nature, 448 (2007), 718-722. |
| [20] | H. Takeuchi, S. Ozawa, N. Ando, C. H. Shih, K. Koyanagi, M. U. Ozawa, et al., Altered p16/MTS1/CDKN2 and cyclin D1/PRAD-1 gene expression is associated with the prognosis of squamous cell carcinoma of the esophagus, Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res., 3 (1997), 2229-2236. |
| [21] | E. S. Antonarakis, AR Signaling in Human Malignancies: Prostate Cancer and Beyond, Cancers, 9 (2017), 7. |
| [22] | T. Ken-Ichi, K. Horie-Inoue, S. Katayama, T. Suzuki, S. Tsutsumi, K. Ikeda, et al., Androgen-responsive long noncoding RNA CTBP1-AS promotes prostate cancer, EMB J., 32 (2013), 1665-1680. |
| [23] | W. Huang, X. Su, W. Yan, Z. Kong, D. Wang, Y. Huang, et al., Overexpression of AR-regulated lncRNA TMPO-AS1 correlates with tumor progression and poor prognosis in prostate cancer, Prostate, 78 (2018), 1248-1261. |
| [24] | M. Yan, H. Qi, J. Li, G. Ye, Y. Shao, T. Li, et al., Identification of SPOP related metabolic pathways in prostate cancer, Oncotarget, 8 (2017), 103032-103046. |
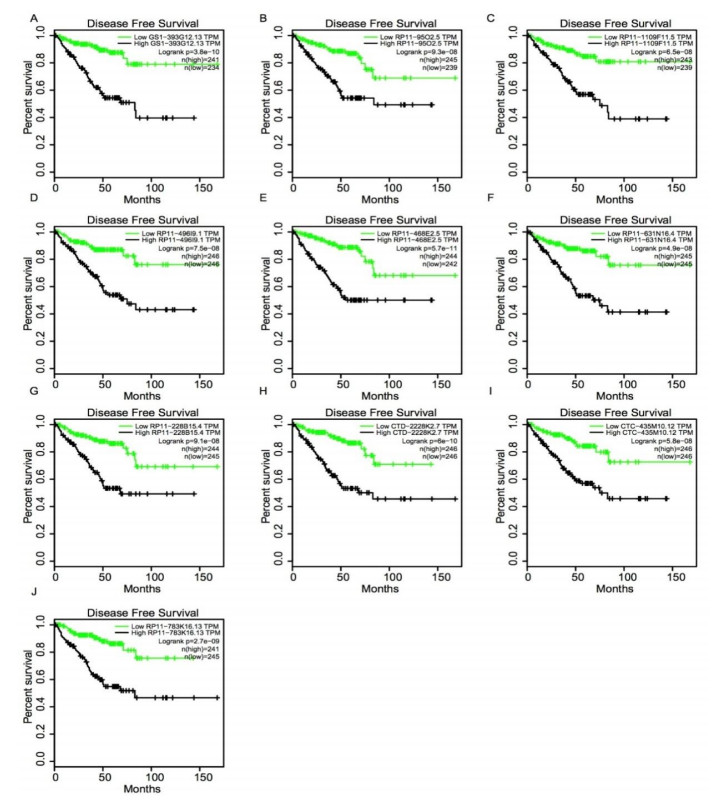
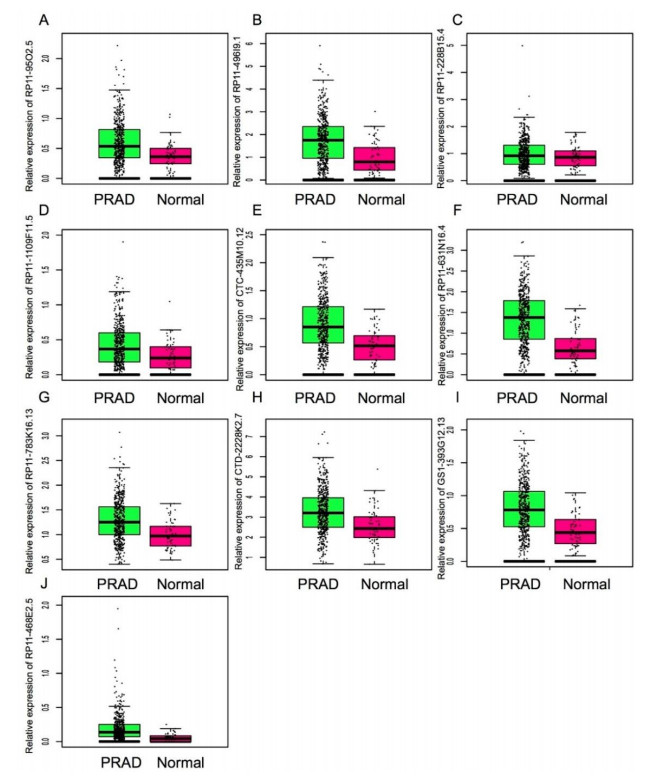
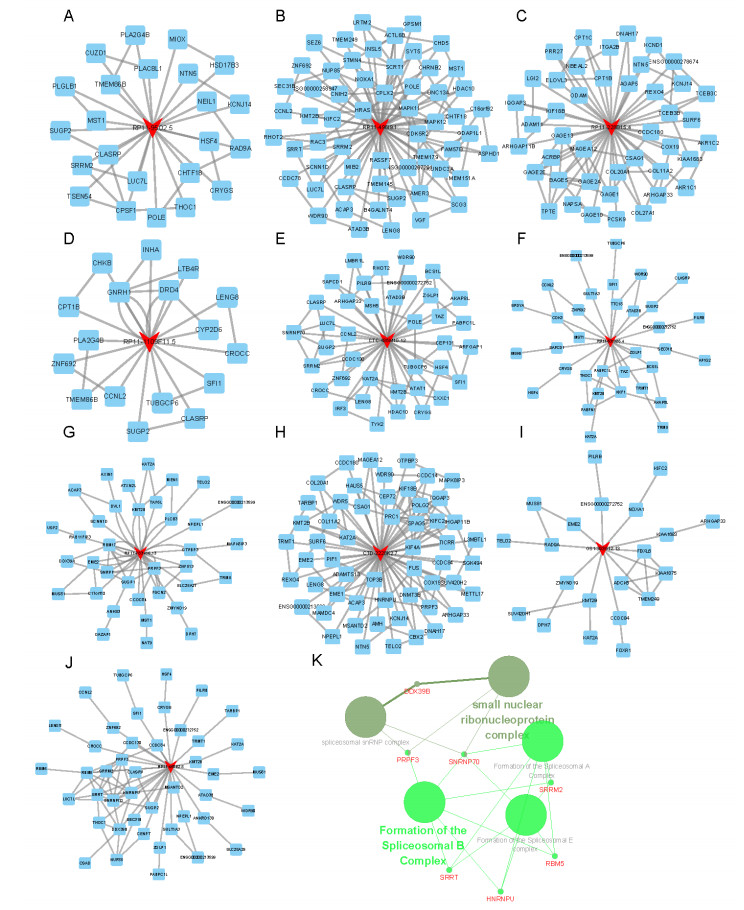

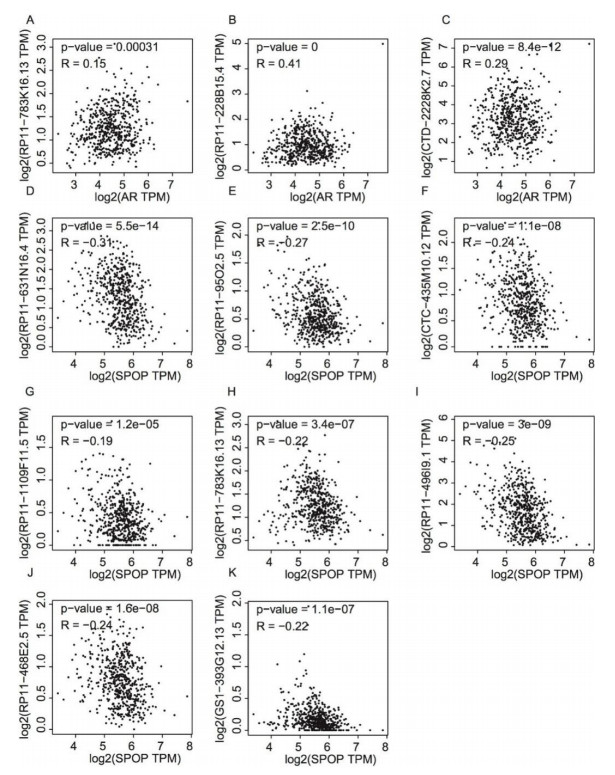
Figures(5)

Ziyu Wu, Sugui Wang, Qiang Li, Qingsong Zhao, Mingming Shao. Identification of 10 differently expressed lncRNAs as prognostic biomarkers for prostate adenocarcinoma[J]. Mathematical Biosciences and Engineering, 2020, 17(3): 2037-2047. doi: 10.3934/mbe.2020108







 DownLoad:
DownLoad: