Citation: Micaela Morettini, Christian Göbl, Alexandra Kautzky-Willer, Giovanni Pacini, Andrea Tura, Laura Burattini. Former gestational diabetes: Mathematical modeling of intravenous glucose tolerance test for the assessment of insulin clearance and its determinants[J]. Mathematical Biosciences and Engineering, 2020, 17(2): 1604-1615. doi: 10.3934/mbe.2020084

| [1] | American Diabetes Association, 2. Classification and diagnosis of diabetes: Standards of medical care in diabetes-2019, Diabetes Care, 42 (2019), S13-S28. |
| [2] | C. Kim, Maternal outcomes and follow-up after gestational diabetes mellitus, Diabetic Med., 31 (2014), 292-301. |
| [3] | B. E. Metzger, T. A. Buchanan, D. R. Coustan, A. de Leiva, D. B. Dunger, D. R. Hadden, et al., Summary and recommendations of the Fifth International Workshop-Conference on Gestational Diabetes Mellitus, Diabetes Care, 30 (2007), S251-S260. |
| [4] | A. Kautzky-Willer, R. Prager, W. Waldhäusl, G. Pacini, K. Thomaseth, O. F Wagner, et al., Pronounced insulin resistance and inadequate β-cell secretion characterize lean gestational diabetes during and after pregnancy, Diabetes Care, 20 (1997), 1717-1723. |
| [5] | C. S. Göbl, L. Bozkurt, T. Prikoszovich, C. Winzer, G. Pacini, A. Kautzky-Willer, Early possible risk factors for overt diabetes after gestational diabetes mellitus, Obstet. Gynecol., 118 (2011), 71-78. |
| [6] | A. Tura, A. Grassi, Y. Winhofer, A. Guolo, G. Pacini, A. Mari, et al., Progression to type 2 diabetes in women with former gestational diabetes: Time trajectories of metabolic parameters, PLoS One, 7 (2012), e50419. |
| [7] | L. Bozkurt, C. S. Göbl, A. Tura, M. Chmelik, T. Prikoszovich, L. Kosi, et al., Fatty liver index predicts further metabolic deteriorations in women with previous gestational diabetes, PLoS One, 7 (2012), e32710. |
| [8] | M. Morettini, C. Castriota, C. Göbl, A. Kautzky-Willer, G. Pacini, L. Burattini, et al., Glucose effectiveness from short insulin-modified IVGTT and its application to the study of women with previous gestational diabetes mellitus, Diabetes Metab. J., in press. |
| [9] | D. C. Polidori, R. N. Bergman, S. T. Chung, A. E. Sumner, Hepatic and extrahepatic insulin clearance are differentially regulated: Results from a novel model-based analysis of intravenous glucose tolerance data, Diabetes, 65 (2016), 1556-1564. |
| [10] | E. Van Cauter, F. Mestrez, J. Sturis, K. S. Polonsky, Estimation of insulin secretion rates from C-peptide levels: Comparison of individual and standard kinetic parameters for C-peptide clearance, Diabetes, 41 (1992), 368-377. |
| [11] | G. Pacini, G. Tonolo, M. Sambataro, M. Maioli, M. Ciccarese, E. Brocco, et al., Insulin sensitivity and glucose effectiveness: Minimal model analysis of regular and insulin-modified FSIGT, Am. J. Physiol. Metab., 274 (2017), E592-E599. |
| [12] | S. E. Kahn, R. L. Prigeon, D. K. McCulloch, E. J. Boyko, R. N. Bergman, M. W. Schwartz, et al., Quantification of the relationship between insulin sensitivity and β-cell function in human subjects: Evidence for a hyperbolic function, Diabetes, 42 (1993), 1663-1672. |
| [13] | A. Tura, A. Mari, T. Prikoszovich, G. Pacini, A. Kautzky-Willer, Value of the intravenous and oral glucose tolerance tests for detecting subtle impairments in insulin sensitivity and beta-cell function in former gestational diabetes, Clin. Endocrinol., 69 (2008), 237-243. |
| [14] | A. Mari, A. Tura, G. Pacini, A. Kautzky-Willer, E. Ferrannini, Relationships between insulinsecretion after intravenous and oral glucose administration in subjects with glucose toleranceranging from normal to overt diabetes, Diabet. Med., 25 (2008), 671-677. |
| [15] | A. Caumo, L. Luzi, First-phase insulin secretion: Does it exist in real life? Considerations on shape and function, Am. J. Physiol. Endocrinol. Metab., 287 (2004), E371-E385. |
| [16] | I. Blumer, E. Hadar, D. R. Hadden, L. Jovanovič, J. H. Mestman, M. H. Murad, et al., Diabetes and pregnancy: An endocrine society clinical practice guideline, J. Clin. Endocrinol. Metab., 98 (2013), 4227-4249. |
| [17] | R. Retnakaran, Y. Qi, C. Ye, A. J. G. Hanley, P. W. Connelly, M. Sermer, et al., Hepatic insulin resistance is an early determinant of declining β-cell function in the first year postpartum after glucose intolerance in pregnancy, Diabetes Care, 34 (2011), 2431-2434. |
| [18] |
S. D. Mittelman, G. W. Van Citters, S. P. Kim, D. A. Davis, M. K. Dea, M. Hamilton-Wessler, et al., Longitudinal compensation for fat-induced insulin resistance includes reduced insulin clearance and enhanced β-cell response, Diabetes, 49 (2000), 2116-2125. doi: 10.2337/diabetes.49.12.2116

|
| [19] | M. Ader, D. Stefanovski, S. P. Kim, J. M. Richey, V. Ionut, K. J. Catalano, et al., Hepatic insulin clearance is the primary determinant of insulin sensitivity in the normal dog, Obesity, 22 (2014), 1238-1245. |
| [20] | M. O. Goodarzi, C. D. Langefeld, A. H. Xiang, Y. I. Chen, X. Guo, A. J. G. Hanley, et al., Insulin sensitivity and insulin clearance are heritable and have strong genetic correlation in mexican americans, Obesity, 22 (2014), 1157-1164. |
| [21] | K. Ohashi, M. Fujii, S. Uda, H. Kubota, H. Komada, K. Sakaguchi, et al., Increase in hepatic and decrease in peripheral insulin clearance characterize abnormal temporal patterns of serum insulin in diabetic subjects, NPJ Syst. Biol. Appl., 4 (2018), 14. |
| [22] | J. Ling, L. Ge, D. H. Zhang, Y. Wang, Z. Xie, J. Tian, et al., DPP-4 inhibitors for the treatment of type 2 diabetes: A methodology overview of systematic reviews, Acta Diabetol., 56 (2019), 7-27. |
| [23] | R. M. Goldenberg, L. Berard, Adding prandial GLP-1 receptor agonists to basal insulin: A promising option for type 2 diabetes therapy, Curr. Med. Res. Opin., 34 (2018), 1-10. |
| [24] | A. Tura, G. Pacini, Y. Yamada, Y. Seino, B. Ahrén, Glucagon and insulin secretion, insulin clearance, and fasting glucose in GIP receptor and GLP-1 receptor knockout mice, Am. J. Physiol. Integr. Comp. Physiol., 316 (2019), R27-R37. |
| [25] | A. Tura, R. Bizzotto, Y. Yamada, Y. Seino, G. Pacini, B. Ahrén, Increased insulin clearance in mice with double deletion of glucagon-like peptide-1 and glucose-dependent insulinotropic polypeptide receptors, Am. J. Physiol. Integr. Comp. Physiol., 314 (2018), R639-R646. |
| [26] | A. Shah, M. M. Holter, F. Rimawi, V. Mark, R. Dutia, J. McGinty, et al., Insulin clearance after oral and intravenous glucose following gastric bypass and gastric banding weight loss, Diabetes Care, 42 (2019), 311-317. |
| [27] | M. Morettini, F. Di Nardo, L. Ingrillini, S. Fioretti, C. Göbl, A. Kautzky-Willer, et al., Glucose effectiveness and its components in relation to body mass index, Eur. J. Clin. Invest., 49 (2019), e13099. |
| [28] | G. Toffolo, R. N. Bergman, D. T. Finegood, C. R. Bowden, C. Cobelli, Quantitative estimation of beta cell sensitivity to glucose in the intact organism. A minimal model of insulin kinetics in the dog, Diabetes, 29 (1980), 979-990. |
| [29] | F. Di Nardo, M. Mengoni, M. Morettini, MATLAB-implemented estimation procedure for model-based assessment of hepatic insulin degradation from standard intravenous glucose tolerance test data, Comput. Methods Programs Biomed., 110 (2012), 215-225. |
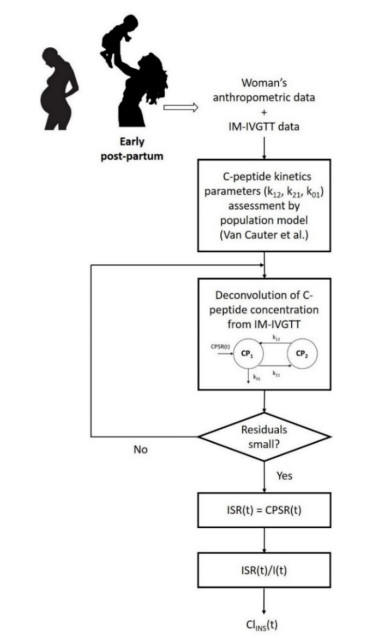
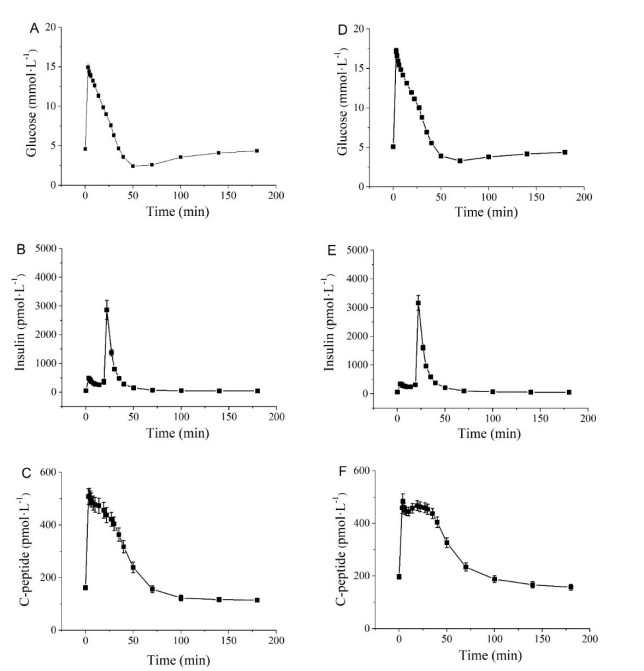
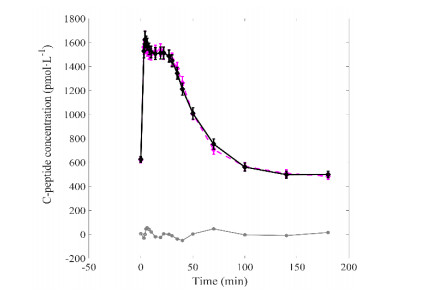
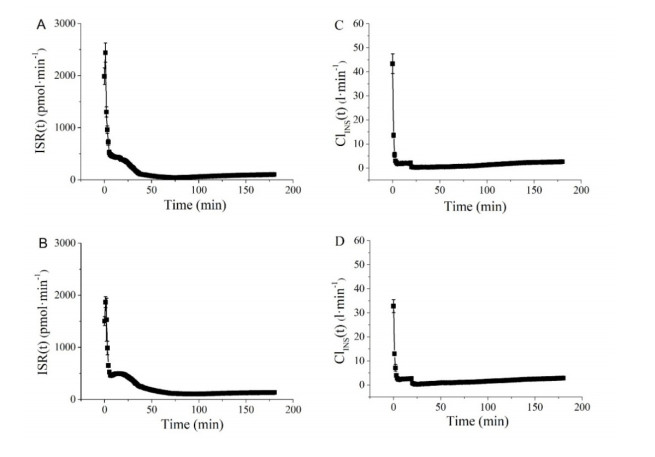
Figures(5) / Tables(2)

Micaela Morettini, Christian Göbl, Alexandra Kautzky-Willer, Giovanni Pacini, Andrea Tura, Laura Burattini. Former gestational diabetes: Mathematical modeling of intravenous glucose tolerance test for the assessment of insulin clearance and its determinants[J]. Mathematical Biosciences and Engineering, 2020, 17(2): 1604-1615. doi: 10.3934/mbe.2020084







 DownLoad:
DownLoad: