Citation: Manikandan Alagumuthu, Divakar Dahiya, Poonam Singh Nigam. Phospholipid—the dynamic structure between living and non-living world; a much obligatory supramolecule for present and future[J]. AIMS Molecular Science, 2019, 6(1): 1-19. doi: 10.3934/molsci.2019.1.1

| [1] | López-Marqués RL, Poulsen LR, Bailly A, et al. (2014) Structure and mechanism of ATP-dependent phospholipid transporters. Biochim biophys Acta 1850: 461–475. |
| [2] |
Lopez-Marques RL, Theorin L, Palmgren MG, et al. (2014) P4-ATPases: lipid flippases in the cell membrane. Pflug Arch Eur J Physiol 466: 1227–1240. doi: 10.1007/s00424-013-1363-4

|
| [3] |
Li J, Wang XL, Zhang T, et al. (2015) A review on phospholipids and their main applications in drug delivery system. Asian J Pharm Sci 10: 81–98. doi: 10.1016/j.ajps.2014.09.004
|
| [4] |
Miao J, Du YZ, Yuan H, et al. (2013) Drug resistance reversal activity of anticancer drug loaded solid lipid nanoparticles in multidrug resistant cancer cells. Colloids Surf B 110: 74–80. doi: 10.1016/j.colsurfb.2013.03.037
|
| [5] | Janben HJ, Steinbuchel A (2014) Fatty acid synthesis in E. coli and its applications towards the production of fatty acid-based biofuels. Biotechnol Biofuels 7: 7. |
| [6] | Hussein J (2013) Cell membrane fatty acids and health. Int J Pharm Pharm Sci 5: 38–46. |
| [7] |
Caforio A, Driessen AJM (2017) Archaeal phospholipids: Structural properties and biosynthesis. Biochim Biophys Acta-Mol Cell Biol Lipids 1862: 1325–1339. doi: 10.1016/j.bbalip.2016.12.006
|
| [8] |
Schiller J, Suss R, Arnhold J, et al. (2004) Matrix-assisted laser desorption and ionization time-of-flight (MALDI-TOF) mass spectrometry in lipid and phospholipid research. Prog Lipid Res 43: 449–488. doi: 10.1016/j.plipres.2004.08.001
|
| [9] |
Irvine RF, Schell MJ (2001) Back in the water: the return of the inositol phosphates. Nat Rev Mol Cell Biol 2: 327–338. doi: 10.1038/35073015
|
| [10] |
Hannun YA, Luberto C, Argraves KM (2001) Enzymes of sphingolipid metabolism: from modular to integrative signaling. Biochemistry 40: 4893–4903. doi: 10.1021/bi002836k
|
| [11] | Cooper GM (2000) The Cell: A Molecular Approach. 2Eds., Sunderland (MA): Sinauer Associates. Available from: https://www.ncbi.nlm.nih.gov/books/NBK9928/. |
| [12] |
Dowhan W, Mileykovskaya E, Bogdanov M (2004) Diversity and versatility of lipid-protein interactions revealed by molecular genetic approaches. Biochim Biophys Acta 1666: 19–39. doi: 10.1016/j.bbamem.2004.04.010
|
| [13] | Adlercreutz P (2000) Enzymatic conversions of glycerophospholipids. In: Bornscheuer, U.T. Editor, Enzymes in Lipid Modification, Weinheim: Wiley, 292–306. |
| [14] |
George CM, Gil-Soo H (2007) Regulation of phospholipid synthesis in Saccharomyces cerevisiae by zinc depletion. Biochim Biophys Acta 1771: 322–330. doi: 10.1016/j.bbalip.2006.05.006
|
| [15] | Perumal Chandran S, Natarajan SB, Senthil Rajan D, et al. (2014) Phospholipids as versatile polymer in drug delivery system. Int J Pharm Pharm Sci 6: 8–11. |
| [16] |
Yang S, Qiao B, Lu SH, et al. (2007) Comparative lipidomics analysis of cellular development and apoptosis in two Taxus cell lines. Biochim Biophys Acta 1771: 600–612. doi: 10.1016/j.bbalip.2007.02.011
|
| [17] |
Mashaghi S, Jadidi T, Koenderink G, et al. (2013) Lipid nanotechnology. Int J Mol Sci 14: 4242–4282. doi: 10.3390/ijms14024242
|
| [18] |
van Hoogevest P, Wendel A (2014) The use of natural and synthetic phospholipids as pharmaceutical excipients. Eur J Lipid Sci Technol 116: 1088–1107. doi: 10.1002/ejlt.201400219
|
| [19] | Burdge GC, Calder PC (2015) Introduction to fatty acids and lipids. World Rev Nutr Diet 112: 1–16. |
| [20] | Berg JM, Tymoczko JL, Stryer L (2002) Biochemistry. 5Eds. New York: W H Freeman. Available from: https://www.ncbi.nlm.nih.gov/books/NBK22581. |
| [21] | Schönfeld P, Reiser G (2013) Why does brain metabolism not favor burning of fatty acids to provide energy? - Reflections on disadvantages of the use of free fatty acids as fuel for brain. J Cerebr Blood F Met 33: 1493–1499. |
| [22] |
Boudière L, Michaud M, Petroutsos D, et al. (2014) Glycerolipids in photosynthesis: composition, synthesis and trafficking. Biochim Biophys Acta-Bioenerg 1837: 470–480. doi: 10.1016/j.bbabio.2013.09.007
|
| [23] |
Siebers M, Brands M, Wewer V, et al. (2016) Lipids in plant-microbe interactions. Biochim Biophys Acta-Mol Cell Biol Lipids 1861: 1379–1395. doi: 10.1016/j.bbalip.2016.02.021
|
| [24] |
Baccile N, Cuvier AS, Prévost S, et al. (2016) Self-assembly mechanism of pH-responsive glycolipids: micelles, fibers, vesicles, and bilayers. Langmuir 32: 10881–10894. doi: 10.1021/acs.langmuir.6b02337
|
| [25] |
Harroun TA, Heller WT, Weiss TM et al. (1999) Theoretical analysis of hydrophobic matching and membrane-mediated interactions in lipid bilayers containing gramicidin. Biophys J 76: 3176–3185. doi: 10.1016/S0006-3495(99)77469-2
|
| [26] |
Peterson BL, Cummings BS (2006) A review of chromatographic methods for the assessment of phospholipids in biological samples. Biomed Chromatogr 20: 227–243. doi: 10.1002/bmc.563
|
| [27] |
Camera E, Picardo M, Presutti C, et al. (2004) Separation and characterization of sphingoceramides by high-performance liquid chromatography–electrospray Ionization mass spectrometry. J Sep Sci 27: 971–976. doi: 10.1002/jssc.200301712
|
| [28] | Chung SY, Moriyama T, Uezu E, et al. (1995) Administration of phosphatidylcholine increases brain acetylcholine concentration and improves memory in mice with dementia. J Nutr 125: 1484–1489. |
| [29] |
Nakamura Y (2017) Plant phospholipid diversity: emerging functions in metabolism and protein-lipid interactions. Trends Plant Sci 22: 1027–1040. doi: 10.1016/j.tplants.2017.09.002
|
| [30] | Bevers EM, Comfurius P, Dekkers DW, et al. (1998) Transmembrane phospholipid distribution in blood cells: control mechanisms and pathophysiological significance. Bio Chem 379: 973–986. |
| [31] |
Sodt AJ, Pastor RW (2014) Molecular modeling of lipid membrane curvature induction by a peptide: more than simply shape. Biophys J 106: 1958–1969. doi: 10.1016/j.bpj.2014.02.037
|
| [32] |
Hama S, Ogino C, Kondo A (2015) Enzymatic synthesis and modification of structures phospholipids: recent advances in enzyme preparation and biocatalytic process. Appl Microbiol Biot 99: 7879–7891. doi: 10.1007/s00253-015-6845-1
|
| [33] |
Fahy E, Subramaniam S, Brown HA, et al. (2005) A comprehensive classification system for lipids. J Lipid Res 46: 839–862. doi: 10.1194/jlr.E400004-JLR200
|
| [34] | Hawthorne JN, Ansell GB (1982) Phospholipids: New Comprehensive Biochemistry. Amsterdam: Elsevier Biomedical Press. |
| [35] | Berg JM, Tymoczko JL, Stryer L (2002) Biochemistry. 5Eds., New York: W H Freeman. Available from: https://www.ncbi.nlm.nih.gov/books/NBK22361/ |
| [36] | Vance DE, Vance JE (2002) Biochemistry of Lipids, Lipoproteins, and Membranes. 4Eds., Amsterdam: Elsevier, 505–526. |
| [37] |
Vance JE (2015) Phospholipid synthesis and transport in mammalian cells. Traffic 16: 1–18. doi: 10.1111/tra.12230
|
| [38] |
Lorant J, Alemayehu G (2010) Importance of the sphingosine base double-bond geometry for the structural and thermodynamic properties of sphingomyelin bilayers. Biophys J 99: 2957–2966. doi: 10.1016/j.bpj.2010.09.020
|
| [39] |
Ramstedt B, Slotte JP (2006) Sphingolipids and the formation of sterol-enriched ordered membrane domains. Biochim Biophys Acta-Biomembr 1758: 1945–1956. doi: 10.1016/j.bbamem.2006.05.020
|
| [40] |
Olsen I, Janzen E (2001) Sphingolipids in bacteria and fungi. Anaerobe 7: 103–112. doi: 10.1006/anae.2001.0376
|
| [41] |
Moskot M, Bocheńska K, Jakóbkiewicz-Banecka J, et al. (2018) Abnormal Sphingolipid World in Inflammation Specific for Lysosomal Storage Diseases and Skin Disorders. Int J Mol Sci 19: 247. doi: 10.3390/ijms19010247
|
| [42] | Helen JS, Sofia KM, Vassilios MK (1989) Lipid composition and structural studies on lipids from the land snail Eobania vermiculata. Naturforsch 44c: 597-608. |
| [43] |
van Hoogevest P, Wendel A (2014) The use of natural and synthetic phospholipids as pharmaceutical excipients. Eur J Lipid Sci Technol 116: 1088–1107. doi: 10.1002/ejlt.201400219
|
| [44] |
Brites P, Waterham HR, Wander, RJA (2004) Functions and biosynthesis of plasmalogens in health and disease. Biochim Biophys Acta 1636: 219–231. doi: 10.1016/j.bbalip.2003.12.010
|
| [45] | Snyder F, Lee TC, Wykle RL (2002) Ether-linked lipids and their bioactive species in biochemistry of lipids, lipoproteins, and membranes. In: Vance, D.E., Vance, J.E. Editors, Biochemistry of Lipids, Lipoproteins and Membranes, 4Eds., Amsterdam: Elsevier, 233–262. |
| [46] |
Li G, Kim J, Huang Z, et al. (2016) Efficient replacement of plasma membrane outer leaflet phospholipids and sphingolipids in cells with exogenous lipids. P Natl Acad Sci USA 113: 14025–14030. doi: 10.1073/pnas.1610705113
|
| [47] |
Montigny C, Lyons J, Champeil P, et al. (2016) On the molecular mechanism of flippase- and scramblase-mediated phospholipid transport. Biochim Biophys Acta 1861: 767–783. doi: 10.1016/j.bbalip.2015.12.020
|
| [48] |
Zhou Y, Wang CO, Cho K, et al. (2015) Signal transduction membrane potential modulates plasma membrane phospholipid dynamics and K-Ras signaling. Science 349: 873–876. doi: 10.1126/science.aaa5619
|
| [49] |
Vance JE (2008) Phosphatidylserine and phosphatidylethanolamine in mammalian cells: two metabolically related amino phospholipids. J Lipid Res 49: 1377–1387. doi: 10.1194/jlr.R700020-JLR200
|
| [50] |
Fadeel B, Xue D (2009) The ins and outs of phospholipid asymmetry in the plasma membrane: roles in health and disease. Criti Rev Biochem Mol Biol 44: 264–277. doi: 10.1080/10409230903193307
|
| [51] |
Suetsugu S, Kurisu S, Takenawa T (2014) Dynamic shaping of cellular membranes by phospholipids and membrane-deforming proteins. Physiol Rev 94: 1219–1248. doi: 10.1152/physrev.00040.2013
|
| [52] | Darland-Ransom M, Wang XC, Sun CL, et al. (2008) Role of C. elegans TAT-1 protein in maintaining plasma membrane phosphatidylserine asymmetry. Science 320: 528–531. |
| [53] |
Mapes J, Chen YZ, Kim A, et al. (2012) CED-1, CED-7, and TTR-52 regulate surface phosphatidylserine expression on apoptotic and phagocytic cells. Curr Biol 22: 1267–1275. doi: 10.1016/j.cub.2012.05.052
|
| [54] |
Helen W (2015) Biological membranes. Essays Biochem 59: 43–69. doi: 10.1042/bse0590043
|
| [55] |
Van Meer G, Voelker DR, Feigenson GW (2008) Membrane lipids: Where they are and how they behave. Nat Rev Mol Cell Biol 9: 112–124. doi: 10.1038/nrm2330
|
| [56] |
Giordano F (2018) Non-vesicular lipid trafficking at the endoplasmic reticulum-mitochondria interface. Biochem Soc Trans 46: 437–452. doi: 10.1042/BST20160185
|
| [57] |
Panatala R, Hennrich H, Holthuis JCM (2015) Inner workings and biological impact of phospholipid flippases. J Cell Sci 128: 2021–2032. doi: 10.1242/jcs.102715
|
| [58] |
Sebastian TT, Baldridge RD, Xu P, et al. (2012) Phospholipid flippases: building asymmetric membranes and transport vesicles. Biochim Biophys Acta 1821: 1068–1077. doi: 10.1016/j.bbalip.2011.12.007
|
| [59] |
Tanaka K, Fujimura KK, Yamamoto T (2011) Functions of phospholipid flippases. J Biochem 149: 131–143. doi: 10.1093/jb/mvq140
|
| [60] |
Contreras FX, Sánchez-Magraner L, Alonso A, et al. (2010) Transbilayer (flip-flop) lipid motion and lipid scrambling in membranes. FEBS Lett 584: 1779–1786. doi: 10.1016/j.febslet.2009.12.049
|
| [61] |
Hankins HM, Baldridge RD, Xu P, et al. (2015) Role of flippases, scramblases and transfer proteins in phosphatidylserine subcellular distribution. Traffic 16: 35–47. doi: 10.1111/tra.12233
|
| [62] |
Lagace TA, Ridgway ND (2013) The role of phospholipids in the biological activity and structure of the endoplasmic reticulum. Biochim Biophys Acta-Mol cell Res 1833: 2499–2510. doi: 10.1016/j.bbamcr.2013.05.018
|
| [63] |
Pomorski TG, Holthuis JCM, Herrmann A, et al. (2004) Tracking down lipid flippases and their biological functions. J Cell Sci 117: 805–813. doi: 10.1242/jcs.01055
|
| [64] |
Nakao H, Ikeda K, Ishihama Y, et al. (2016) Membrane-spanning sequences in endoplasmic reticulum proteins promote phospholipid flip-flop. Biophys J 110: 2689–2697. doi: 10.1016/j.bpj.2016.05.023
|
| [65] |
Montigny C, Lyons J, Champeil P, et al. (2016) On the molecular mechanism of flippase- and scramblase-mediated phospholipid transport. Biochim Biophys Acta-Mol Cell Biol Lipids 1861: 767–783. doi: 10.1016/j.bbalip.2015.12.020
|
| [66] |
Siarheyeva A, Sharom FJ (2009) The ABC transporter MsbA interacts with lipid A and amphipathic drugs at different. Biochem J 419: 317–328. doi: 10.1042/BJ20081364
|
| [67] |
Casadei MA, Mañas P, Niven G, et al. (2002) Role of membrane fluidity in pressure resistance of Escherichia coli NCTC 8164. Appl Environ Microbiol 68: 5965–5972. doi: 10.1128/AEM.68.12.5965-5972.2002
|
| [68] |
Sajbidor J (1997) Effect of some environmental factors on the content and composition of microbial membrane lipids. Crit Rev Biotechnol 17: 87–103. doi: 10.3109/07388559709146608
|
| [69] |
Goldstein DB (1984) The effects of drugs on membrane fluidity. Annu Rev Pharmacol Toxicol 24: 43–64. doi: 10.1146/annurev.pa.24.040184.000355
|
| [70] | Begley TP, Guschina IA, Harwood JL (2008) Lipids: chemical diversity. In: Begley, T.P. Editor, Wiley Encyclopedia of Chemical Biology. |
| [71] |
Yagüe G, Segovia M, Valero-Guillén PL (1997) Acyl phosphatidylglycerol: a major phospholipid of Corynebacterium amycolatum. FEMS Microbiol Lett 151: 125–130. doi: 10.1016/S0378-1097(97)00137-7
|
| [72] |
Mazzella N, Molinet J, Syakti AD, et al. (2004) Bacterial phospholipid molecular species analysis by ion-pair reversed-phase HPLC/ESI/MS. J Lipid Res 45: 1355–1363. doi: 10.1194/jlr.D300040-JLR200
|
| [73] | Niepel MTH, Wray V, Abraham WR (2006) Intraspecific variation of unusual phospholipids from Corynebacterium spp. containing a Novel Fatty acid. J Bacteriol 180: 4650–4657. |
| [74] | Pluschke G, Hirota Y, Overath P (1978) Function of phospholipids in Escherichia coli. characterization of a mutant deficient in cardiolipin synthesis. J Biol Chem 253: 5048–5055. |
| [75] |
Albelo ST, Domenech CE (1997) Carbons from choline present in the phospholipids of Pseudomonas aeruginosa. FEMS microbiol lett 156: 271–274. doi: 10.1111/j.1574-6968.1997.tb12739.x
|
| [76] |
Wang XG, Scagliotti JP, Hu LT (2004) Phospholipid synthesis in Borrelia burgdorferi BB0249 and BB0721 encode functional phosphatidylcholine synthase phosphatidylglycerol phosphate synthase proteins. Microbiol 150: 391–397. doi: 10.1099/mic.0.26752-0
|
| [77] |
Burgdorfer W, Barbour AG, Hayes SF, et al. (1982) Lyme disease-a tick-borne spirochetosis? Science 216: 1317–1319. doi: 10.1126/science.7043737
|
| [78] | Oliver JD, Colwell R (1973) Extractable lipids of Gram-negative marine bacteria: phospholipid composition. J Bacteriol 114: 897–908. |
| [79] | Yakimov MM, Golyshin PN, Lang S, et al. (1998) Alcanivorax borkurnensis gen. now., sp. nov., a new, hydrocarbon-degrading and surfactant-producing marine bacterium. Int J Sys Bacteriol 48: 339–348. |
| [80] |
Rottet S, Besagni C, Kessler F (2015) The role of plastoglobules in thylakoid lipid remodeling during plant development. Biochim Biophys Acta-Bioenerg 1847: 889–899. doi: 10.1016/j.bbabio.2015.02.002
|
| [81] |
Gao QM, Yu K, Xia Y, Shine MB, et al. (2014) Mono- and digalactosyldiacylglycerol lipids function non-redundantly to regulate systemic acquired resistance in plants. Cell Rep 9: 1681–1691. doi: 10.1016/j.celrep.2014.10.069
|
| [82] |
Dubots E, Botté C, Boudière L, et al. (2012) Role of phosphatidic acid in plant galactolipid synthesis. Biochimie 94: 86–93. doi: 10.1016/j.biochi.2011.03.012
|
| [83] |
Kobayashi K, Awai K, Nakamura M, et al. (2009) Type-B Monogalactosyl diacylglycerol synthases are involved in phosphate starvation-induced lipid remodeling, and are crucial for low-phosphate adaptation. Plant J 57: 322–331. doi: 10.1111/j.1365-313X.2008.03692.x
|
| [84] |
Lassègue B, Alexander RW, Clark M, et al. (1993) Phosphatidylcholine is a major source of phosphatidic acid and diacylglycerol in angiotensin II-stimulated vascular smooth-muscle cells. Biochem J 292: 509–517. doi: 10.1042/bj2920509
|
| [85] |
Harwood JL, Nicholls RG (1979) The plant sulpholipid-a major component of the sulphur cycle. Biochem Soc T 7: 440–447. doi: 10.1042/bst0070440
|
| [86] |
Joyard J, Teyssier E, Miege C, et al. (1998) The biochemical machinery of plastid envelope membranes. Plant Physiol 118: 715–723. doi: 10.1104/pp.118.3.715
|
| [87] |
Damnjanovic J, Iwasaki Y (2013) Phospholipase D as a catalyst: application in phospholipid synthesis, molecular structure, and protein engineering. J Biosci Bioeng 116: 271–280. doi: 10.1016/j.jbiosc.2013.03.008
|
| [88] |
Zhao P (2015) Phospholipase D and phosphatidic acid in plant defense response: from protein-protein and lipid-protein interaction to hormone signaling. J Exp Bot 66: 1721–1736. doi: 10.1093/jxb/eru540
|
| [89] | Bodin S, Giuriato S, Ragab J, et al. (2001) Production of phosphatidylinositol 3,4,5-trisphosphate and phosphatidic acid in platelet rafts: evidence for a critical role of cholesterol enriched domains in human platelet activation. Biochemistry 40, 50: 15290–15299. |
| [90] |
Meijer HG, Munnik T (2003) Phospholipid-based signaling in plants. Annu Rev Plant Biol 54: 265–306. doi: 10.1146/annurev.arplant.54.031902.134748
|
| [91] |
Takashi T, Thomas L (2017) Intra-mitochondrial phospholipid trafficking. Biochim Biophys Acta-Mol Cell Biol Lipids 1862: 81–89. doi: 10.1016/j.bbalip.2016.08.006
|
| [92] |
Cowan KA (2006) Phospholipids as plant growth regulators. J Plant Growth Regul 48: 97–109. doi: 10.1007/s10725-005-5481-7
|
| [93] |
Munnik T (2001) Phosphatidic acid: an emerging plant lipid second messenger. Trends Plant Sci 6: 227–233. doi: 10.1016/S1360-1385(01)01918-5
|
| [94] |
Testerink C, Munnik T (2005) Phosphatidic acid: a multifunctional stress signaling lipid in plants. Trends Plant Sci 10: 368–375. doi: 10.1016/j.tplants.2005.06.002
|
| [95] | Lodish H, Berk A, Zipursky SL, et al. (2000) Molecular cell Biology. 4Eds., New York: W. H. Freeman. Available from: https://www.ncbi.nlm.nih.gov/books/NBK21583/. |
| [96] |
Penno A, Hackenbroich G, Thiele C (2013) Phospholipids and lipid droplets. Biochim Biophys Acta-Mol Cell Biol Lipids 1831: 589–594. doi: 10.1016/j.bbalip.2012.12.001
|
| [97] |
Gupta G, Surolia A (2010) Glycosphingolipids in microdomain formation and their spatial organization. FEBS Lett 584: 1634–1641. doi: 10.1016/j.febslet.2009.11.070
|
| [98] | Alberts B, Johnson A, Lewis J (2002) Molecular biology of the cell. 4Eds., NewYork: Garland Science. |
| [99] |
Dufourc EJ (2008) Sterols and membrane dynamics. J Chem Biol 1: 63–77. doi: 10.1007/s12154-008-0010-6
|
| [100] |
Singer SJ (2004) Some early history of membrane molecular biology. Annu Rev Physiol 66: 1–27. doi: 10.1146/annurev.physiol.66.032902.131835
|
| [101] |
Stoeckenius W, Engelman DM (1969) Current models for the structure of biological membranes. J Cell Biol 42: 613–646. doi: 10.1083/jcb.42.3.613
|
| [102] | Simons K, Sampaio JL (2011) Membrane organization and lipid rafts. Cold Spring Harb Perspect Biol 3: a004697. |
| [103] |
Kim SH, Song HE, Kim SJ, et al. (2017) Quantitative structural characterization of phosphatidylinositol phosphates from biological samples. J Lipid Res 58: 469–478. doi: 10.1194/jlr.D069989
|
| [104] |
Kim Y, Shanta SR, Zhou LH, et al. (2010) Mass spectrometry-based cellular phosphoinositides profiling and phospholipid analysis: a brief review. Exp Mol Med 42: 1–11. doi: 10.3858/emm.2010.42.1.001
|
| [105] |
Berridge MJ (1993) Inositol trisphosphate and calcium signaling. Nature 361: 315–325. doi: 10.1038/361315a0
|
| [106] | Pronk JT, De BJC, Bos P (1992) Anaerobic growth of Thiobacillus ferrooxidans. Appl Environ Microbiol 58: 2227–2230. |
| [107] |
Rawlings DE (2002) Heavy metal mining using microbes. Annu Rev Microbiol 56: 65–91. doi: 10.1146/annurev.micro.56.012302.161052
|
| [108] |
Rampelotto PH (2010) Resistance of microorganisms to extreme environmental conditions and its contribution to astrobiology. Sustainability 2: 1602–1623. doi: 10.3390/su2061602
|
| [109] |
Haruta S, Kanno N (2015) Survivability of microbes in natural environments and their ecological impacts. Microbes Environ 30: 123–125. doi: 10.1264/jsme2.ME3002rh
|
| [110] | Bloem J, de Ruiter P, Bouwman LA (1997) Soil Food webs and nutrient cycling in agroecosystems. In: van Elsas, JD., Trevors, J.T., Willington, E. Editors, Modern Soil Microbiology. New York: Marcel Dekker, 245–278. |
| [111] | Kaur A, Kaur A, Choudhary R, et al. (2005) Phospholipid fatty acid-a bioindicator of environment monitoring and assessment in soil ecosystem. Curr Sci 89: 1103–1112. |
| [112] | Hosokawa M, Minami K, Kohno H, et al. (1999) Differentiation- and apoptosis-inducing activities of phospholipids containing docosahexaenoic acid for mouse myeloid leukemia M1 cells. Fish Sci 65: 789–799. |
| [113] |
Chaurio RA, Janko C, Muñoz LE, et al. (2009) Phospholipids: key players in apoptosis and immune regulation. Molecules 14: 4892–4914. doi: 10.3390/molecules14124892
|
| [114] | Hoy CE, Xu X (2001) Structured triacylglycerols. In: Gunstone FD. Editor, Structured and Modified Lipid. New York: Marcel Dekker, 209–240. |
| [115] |
Willers C, Jansen VRPJ, Claassens S (2015) Phospholipid fatty acid profiling of microbial communities-a review of interpretations and recent applications. J App Microbiol 119: 1207–1218. doi: 10.1111/jam.12902
|
| [116] | Ashraf MZ, Kar NS, Podrez EA (2009) Oxidized phospholipids: biomarker for cardiovascular diseases. Int J Biochem Cell B 4: 1241–1244. |
| [117] |
van Hoogevest P, Wendel A (2014) The use of natural and synthetic phospholipids as pharmaceutical excipients. Eur J Lipid Sci Tech 116: 1088–1107. doi: 10.1002/ejlt.201400219
|
| [118] | Diehl BWK, Ockels W (1995) Phospholipids: characterization, metabolism, and novel biological applications. In: Cevc, G., Paltauf, F. Editors, Proceedings of the 6th International Colloquium, USA: AOCS Press, 29–32. |
| [119] |
Descalzo AM, Insani EM, Pense NA (2003) Light-scattering detection of phospholipids resolved by HPLC. Lipids 38: 999–1003. doi: 10.1007/s11745-003-1154-1
|
| [120] | Campbell NA, Williamson B, Heyden RJ (2015) Biology Exploring Life. Boston, Massachusetts: Pearson Prentice Hall. |
| [121] |
Zachowski A (1993) Phospholipids in animal eukaryotic membranes: transverse asymmetry and movement. Biochem J 294: 1–14. doi: 10.1042/bj2940001
|
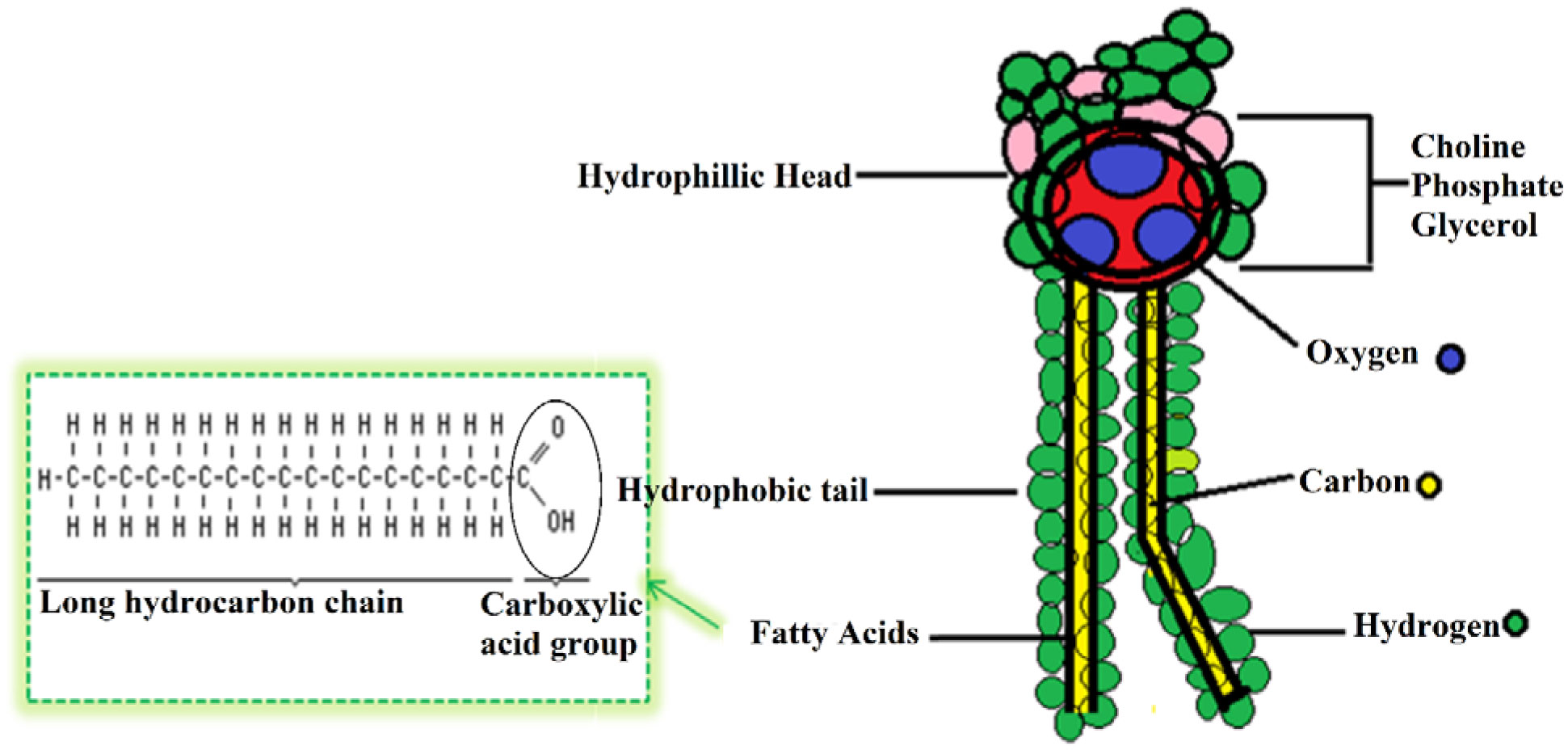
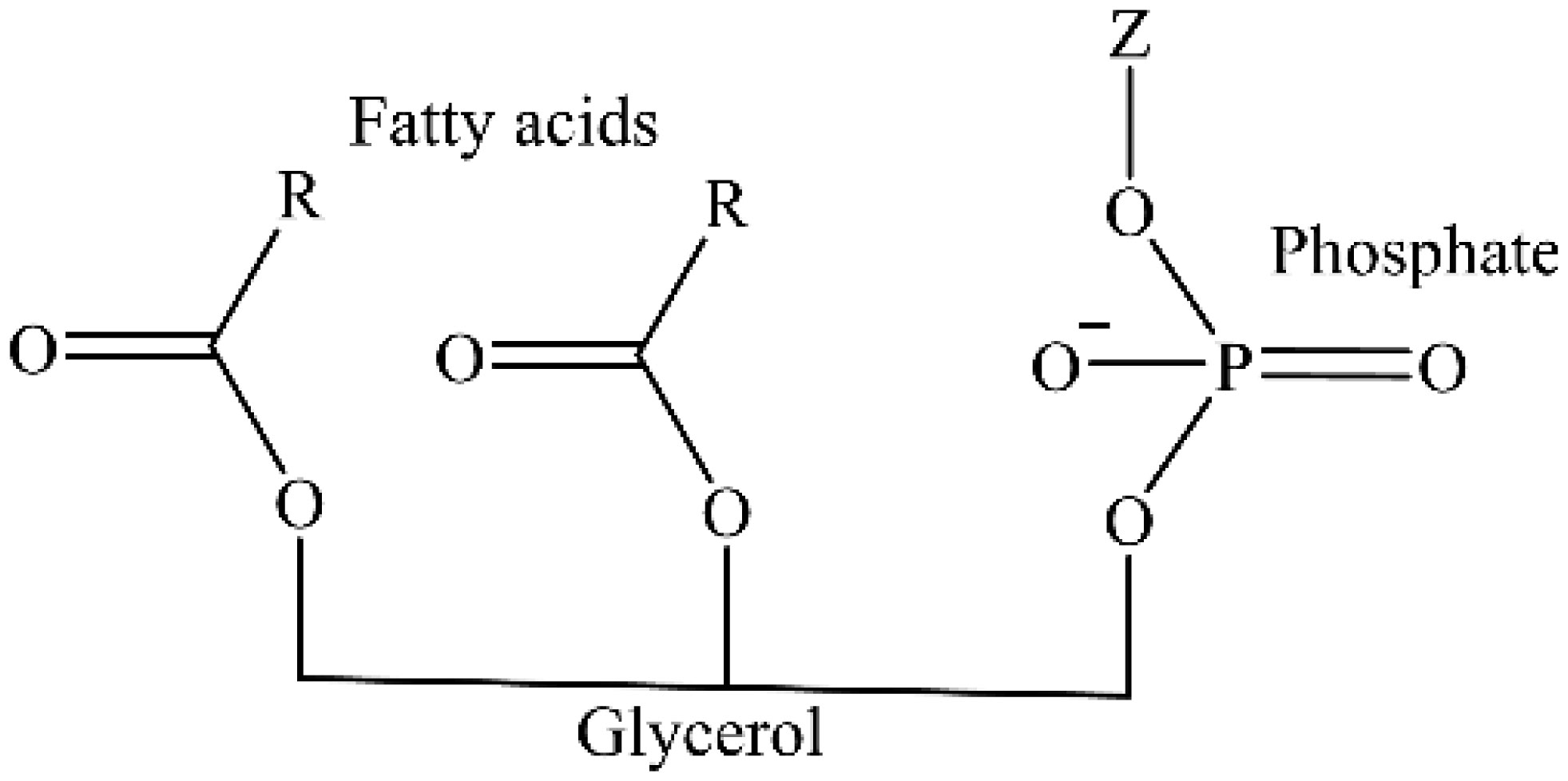
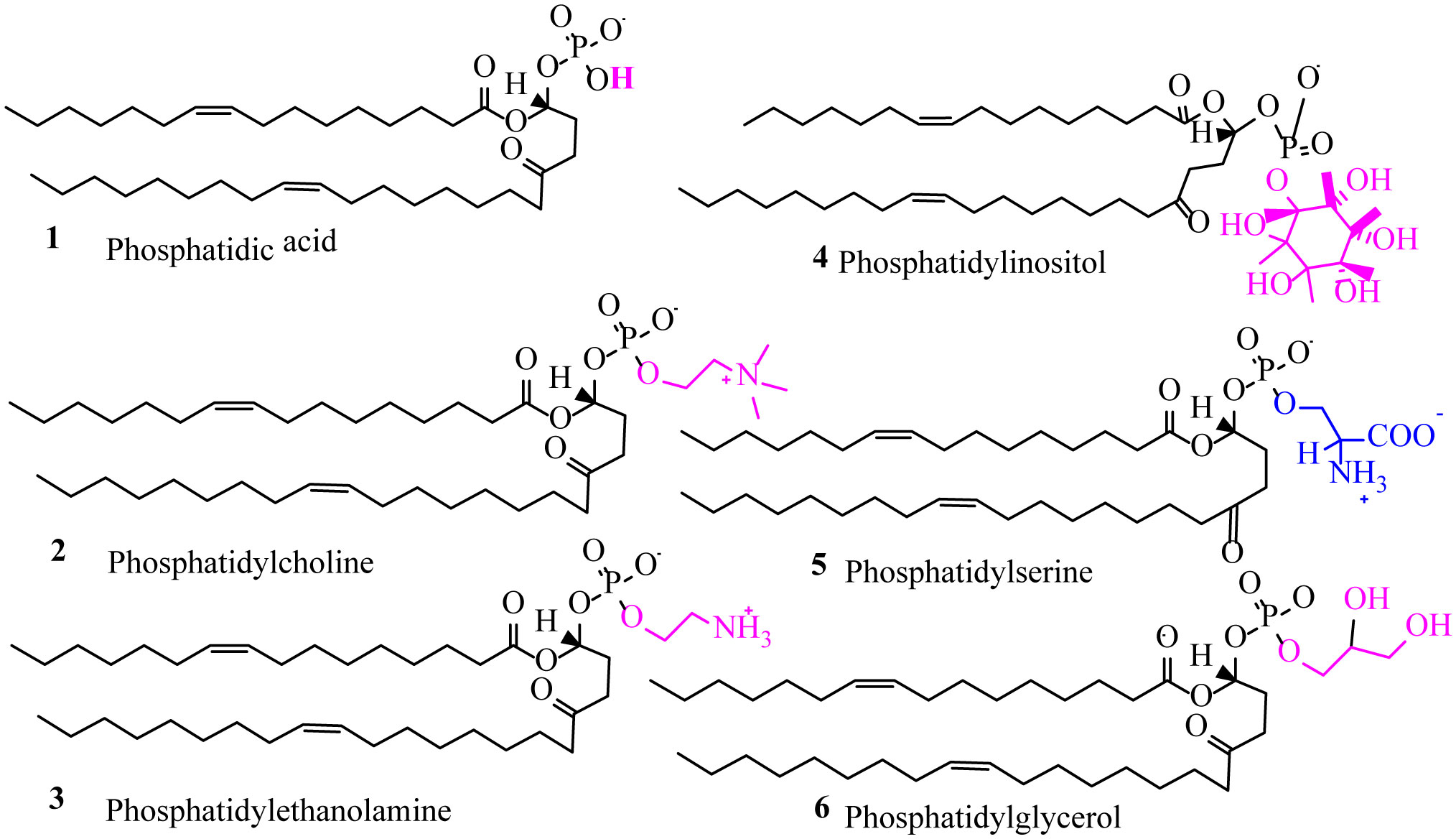
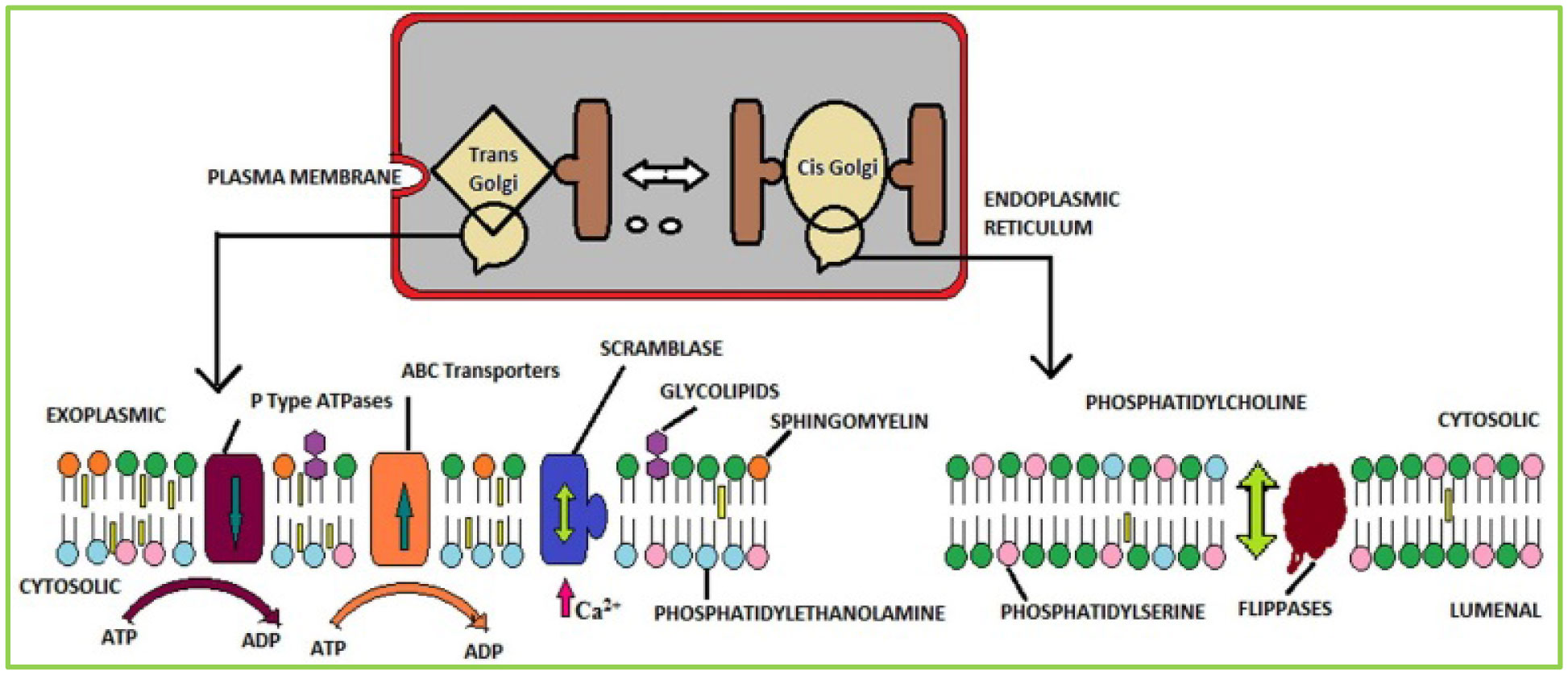
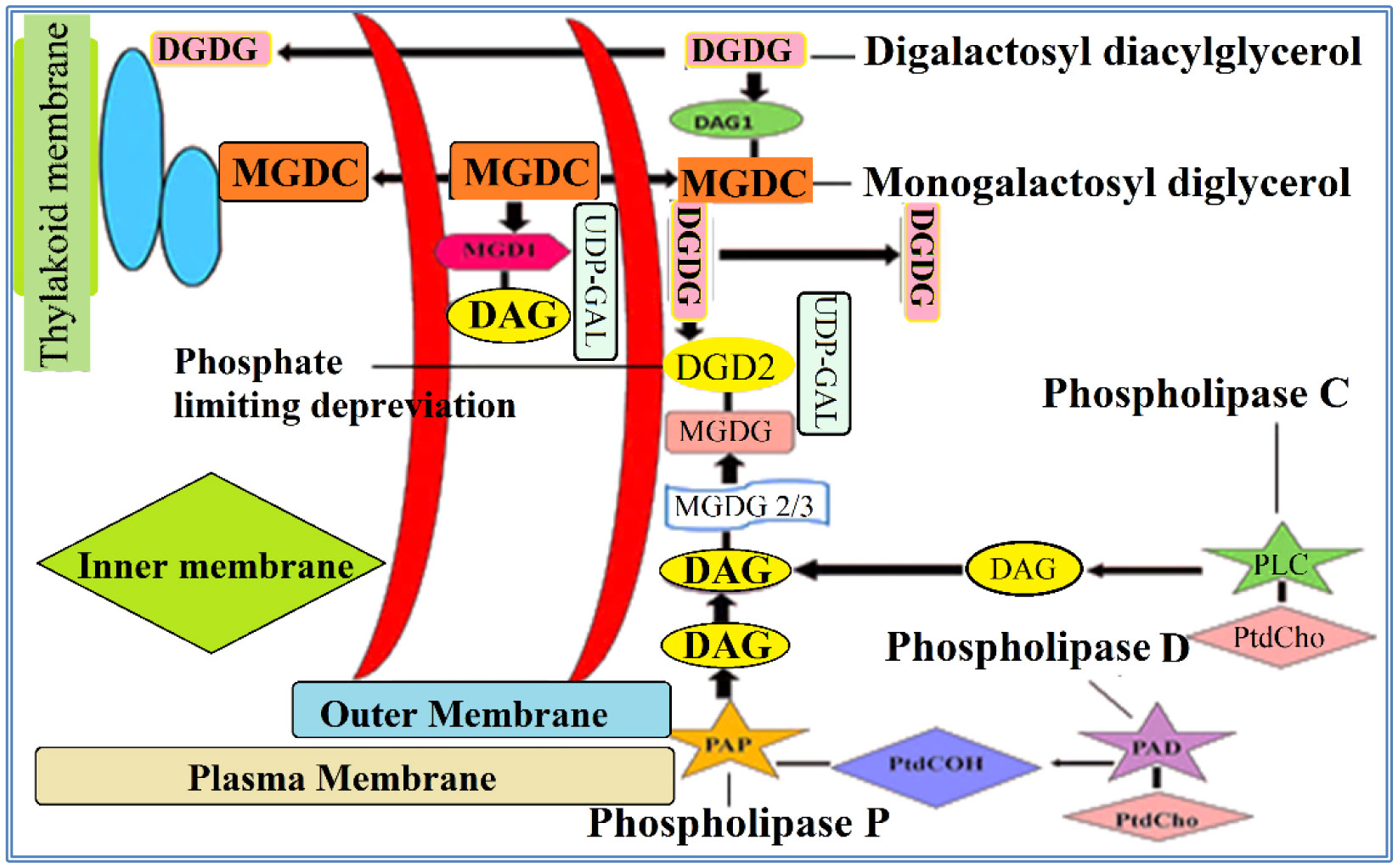
Figures(5) / Tables(3)

Manikandan Alagumuthu, Divakar Dahiya, Poonam Singh Nigam. Phospholipid—the dynamic structure between living and non-living world; a much obligatory supramolecule for present and future[J]. AIMS Molecular Science, 2019, 6(1): 1-19. doi: 10.3934/molsci.2019.1.1







 DownLoad:
DownLoad: