Citation: Bo Sun, Ming Wei, Wei Wu, Binbin Jing. A novel group decision making method for airport operational risk management[J]. Mathematical Biosciences and Engineering, 2020, 17(3): 2402-2417. doi: 10.3934/mbe.2020130

| [1] | F. Netjasov, M. Janic, A review of research on risk and safety modeling in civil aviation, J. Air Transp. Manag., 14 (2008), 213-220. |
| [2] | M. Rong, M. Luo, Y. Chen, The research of airport operational risk alerting model, Dig. Hum. Model., 9745 (2016), 586-595. |
| [3] | F. Fabio, M. Massimo, T. Riccardo, Electrical safety of aeronautical ground lighting systems, IEEE Trans. Ind. Appl., 51 (2015), 2003-2008. |
| [4] | K. T. Atanassov, Intuitionistic fuzzy sets, Fuzzy Sets Syst., 75 (1995), 401-402. |
| [5] | A. Kaur, A. Kumar, S. S. Appadoo, A note on approaches to interval intuitionistic trapezoidal fuzzy multiple attribute decision making with incomplete weight information, Int. J. Fuzzy Syst., 17 (2019), 484-489. |
| [6] | A. Si, S. Das, S. Kar, An approach to rank picture fuzzy nnumbers for decision making problems, Decis. Mak. Appl. Manag. Eng., 2 (2019), 54-64. |
| [7] | K. Q. Zhao, Set pair analysis and its primary application, Explor. Nat., 13 (1994), 67-71. |
| [8] | Q. Zou, J. Zhou, C. Zhou, Comprehensive flood risk assessment based on set pair analysis-variable fuzzy sets model and fuzzy AHP, Stoch. Environ. Res. Risk Assess., 27 (2013), 525-546. |
| [9] | F. Yan, K. Xu, D. Li, et al., A novel hazard assessment method for biomass gasification stations based on extended set pair analysis, Plos One, 12 (2017), 1-17. |
| [10] | Y. S. Pang, J. K. Liu, L. Zhang, Research on government investment group decision-making method for mega construction projects based on intuitionistic fuzzy distance, J. Xi'an Univ. Arch. Tech., 47 (2015), 734-739. |
| [11] | C. Q. Tan, Q. Zhang, Aggregation of opinion in group decision making based on intuitionistic fuzzy distances, Math. Prac. Theo., 36 (20062), 119-124. |
| [12] | G. Li, G. Kou, Y. Peng, A group decision making model for integrating heterogeneous information, IEEE Trans. Syst., Man Cybern., 6 (2018), 982-992. |
| [13] | H. H. Zhang, G. Kou, L. Peng, Soft consensus cost models for group decision making and economic interpretations, Eur. J. Opera. Res., 3 (2019), 964-980. |
| [14] | F. Liu, G.Aiwu,V. Lukovac, M. Vukić, A multicriteria model for the selection of the transport service provider: A single valued neutrosophic DEMATEL multicriteria model, Dec. Mak., 1 (2018), 121-130. |
| [15] | L. Shi, R. Luo, Research on risk early-warning model in airport flight area based on information entropy attribute reduction and BP neural network, Int. J. Sec. Its Appl., 9 (2015), 313-322. |
| [16] | H. B. Jin, Y. Hong, Y. M. Cai, Safety evaluation model of airport operation based on factor analysis, Ind. Safety Environ. Prot., 41 (2015), 62-65. |
| [17] | Y. H. Chang, H. H. Yng, Y. J. Hsiao, Human risk factors associated with pilots in runway excursions, Acc. Anal. Prev., 94 (2016), 227-237. |
| [18] | F. Hofer, O. E. Wetter, Operational and human factors issues of new airport security technology-two case studies, J. Transp. Sec., 5 (2012), 21-29. |
| [19] | X. Li, F. Li, X. Chen, Study on the configuration and capacity of the lateral runway based on the airport green operation, IOP Confer. Series Eart. Environ. Sci., 63 (2017), 12-32. |
| [20] | M. Mostafaee, P. Nassiri, M. H. Behzadi, Investigation of noise pollution in ground safety section of mehrabad airport and its relation with employees hearing loss, J. Health Safety Work, 5 (2015), 7-8. |
| [21] | X. L. Luo, D. X. Chen, Research on risk evaluation using rnp technology for operation into high elevation airports with critical terrain, 2nd Int. Sympos. Air. Airworth. (ISAA), 17 (2011), 125-140. |
| [22] | D. G. Sun, J. Sun, M. Wang, Application of the improved bow-tie analysis technology in civil airport safey, J. Safety Sci. Tech., 4 (2010), 85-90. |
| [23] | H. W. Liu, F. Zhang, Y. M. Cai, Risk assessment of the airport operation based on entropy weight, J. Chongqing Univ. Tech., 30 (2016), 177-184. |
| [24] | M. J. Rezaee, S. Yousefi, An Intelligent decision-making approach for identifying and analyzing airport risks, J. Air Transp. Manag., 68 (2017), 1-14. |
| [25] |
D. K. Y. Wong, D. E. Pitfield, R. E. Caves, et al., The development of a more risk-sensitive and flexible airport safety area strategy: Part I, the development of an improved accident frequency model, Safety Sci., 47 (2009), 913-924. doi: 10.1016/j.ssci.2008.09.011

|
| [26] |
X. Nie, R. Batta, C. G. Drury, et al., Passenger grouping with risk levels in an airport security system, Eur. J. Opera. Res., 194 (2009), 574-584. doi: 10.1016/j.ejor.2007.12.027
|
| [27] | M. Hadjimichael, A fuzzy expert system for aviation risk assessment, Expert Syst. Appl., 36 (2009), 6512-6519. |
| [28] | X. Qin, F. Luo, On the safety risk assessment of the airport flight area based on the catas-trophe theory and fuzzy set, J. Safety. Environ., 18 (2018), 1730-1736. |
| [29] | X. X. Tang, F. Luo, Human risk assessment of the airport runway incursion based on the grey clustering, J. Univ. Elect. Sci. Tech. Cha., 328 (2015), 11-20. |
| [30] | S. Chakraborty, N. Creutzfeldt-Banda, 2010 meltdown-airport closure risk communications in London and NYC. Eur. J. Risk Reg., 2 (2011), 108-110. |
| [31] | D. Doherty, Operational safety implications on airfield services at a civilian airport, IEEE Semi. Air. Elec. Syst. Mtg. Safety Clg., (2000), 85-94. |
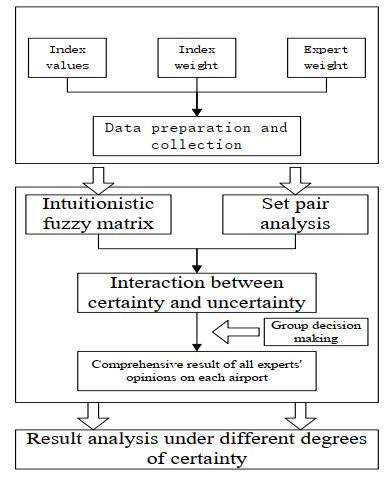
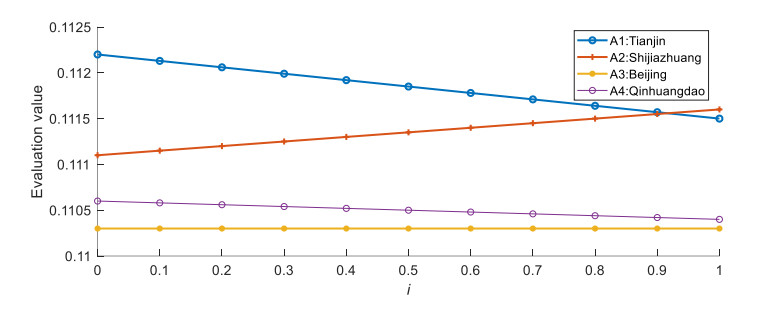
Figures(2) / Tables(5)

Bo Sun, Ming Wei, Wei Wu, Binbin Jing. A novel group decision making method for airport operational risk management[J]. Mathematical Biosciences and Engineering, 2020, 17(3): 2402-2417. doi: 10.3934/mbe.2020130







 DownLoad:
DownLoad: