Citation: Yusuke Morita, Mai Okumura, Issay Narumi, Hiromi Nishida. Sensitivity of Deinococcus grandis rodZ deletion mutant to calcium ions results in enhanced spheroplast size[J]. AIMS Microbiology, 2019, 5(2): 176-185. doi: 10.3934/microbiol.2019.2.176

| [1] |
Morita Y, Nishida H (2018) The common ancestor of Deinococcus species was rod-shaped. Open Bioinfo J 11: 252–258. doi: 10.2174/1875036201811010252

|
| [2] |
Gupta RS (2011) Origin of diderm (Gram-negative) bacteria: antibiotic selection pressure rather than endosymbiosis likely led to the evolution of bacterial cells with two membranes. Antonie van Leeuwenhoek 100: 171–182. doi: 10.1007/s10482-011-9616-8
|
| [3] |
Raetz CR, Whitfield C (2002) Lipopolysaccharide endotoxins. Annu Rev Biochem 71: 635–700. doi: 10.1146/annurev.biochem.71.110601.135414
|
| [4] |
Farci D, Bowler MW, Kirkpatrick J, et al. (2014) New features of the cell wall of the radio-resistant bacterium Deinococcus radiodurans. Biochim Biophys Acta 1838: 1978–1984. doi: 10.1016/j.bbamem.2014.02.014
|
| [5] |
Tian B, Wang H, Ma X, et al. (2010) Proteomic analysis of membrane proteins from a radioresistant and moderate thermophilic bacterium Deinococcus geothermalis. Mol BioSyst 6: 2068–2077. doi: 10.1039/c004875e
|
| [6] | Oyaizu H, Stackebrandt E, Schleifer KH, et al. (1987) A radiation-resistant rod-shaped bacterium, Deinobacter grandis gen. nov., sp. nov., with peptidoglycan containing ornithine. Int J Syst Bacteriol 37: 62–67. |
| [7] |
Nishino K, Morita Y, Takahashi S, et al. (2018) Enlargement of Deinococcus grandis spheroplasts requires Mg2+ or Ca2+. Microbiology 164: 1361–1371. doi: 10.1099/mic.0.000716
|
| [8] | Nishino K, Nishida H (2019) Calcium ion induces outer membrane fusion of Deinococcus grandis spheroplasts to generate giant spheroplasts with multiple cytoplasms. FEMS Microbiol Lett 366: fny282. |
| [9] |
Nishino K, Tsuchikado R, Nishida H (2019) Sugar enhances outer membrane fusion in Deinococcus grandis spheroplasts to generate calcium ion-dependent extra-huge cells. FEMS Microbiol Lett 366: fnz087. doi: 10.1093/femsle/fnz087
|
| [10] | Anderson R, Hansen K (1985) Structure of a novel phosphoglycolipid from Deinococcus radiodurans. J Biol Chem 260: 12219–12223. |
| [11] | Huang Y, Anderson R (1989) Structure of a novel glucosamine-containing phosphoglycolipid from Deinococcus radiodurans. J Biol Chem 264: 18667–18672. |
| [12] |
Makarova KS, Aravind L, Wolf YI, et al. (2001) Genome of the extremely radiation-resistant bacterium Deinococcus radiodurans viewed from the perspective of comparative genomics. Microbiol Mol Biol Rev 65: 44–79. doi: 10.1128/MMBR.65.1.44-79.2001
|
| [13] |
Bos MP, Robert V, Tommassen J (2007) Biogenesis of the gram-negative bacterial outer membrane. Annu Rev Microbiol 61: 191–214. doi: 10.1146/annurev.micro.61.080706.093245
|
| [14] |
Bendezú FO, Hale CA, Bernhardt TG, et al. (2009) RodZ (YfgA) is required for proper assembly of the MreB actin cytoskeleton and cell shape in E. coli. EMBO J 28: 193–204. doi: 10.1038/emboj.2008.264
|
| [15] |
Shiomi D, Sakai M, Niki H (2008) Determination of bacterial rod shape by a novel cytoskeletal membrane protein. EMBO J 27: 3081–3091. doi: 10.1038/emboj.2008.234
|
| [16] |
Alyahya SA, Alexander R, Costa T, et al. (2009) RodZ, a component of the bacterial core morphogenic apparatus. Proc Natl Acad Sci USA 106: 1239–1244. doi: 10.1073/pnas.0810794106
|
| [17] |
van den Ent F, Johnson CM, Persons L, et al. (2010) Bacterial actin MreB assembles in complex with cell shape protein RodZ. EMBO J 29: 1081–1090. doi: 10.1038/emboj.2010.9
|
| [18] |
Jones LJ, Carballido-Lopez R, Errington J (2001) Control of cell shape in bacteria: helical, actin-like filaments in Bacillus subtilis. Cell 104: 913–922. doi: 10.1016/S0092-8674(01)00287-2
|
| [19] |
van den Ent F, Amos LA, Lowe J (2001) Prokaryotic origin of the actin cytoskeleton. Nature 413: 39–44. doi: 10.1038/35092500
|
| [20] |
Funayama T, Narumi I, Kikuchi M, et al. (1999) Identification and disruption analysis of the recN gene in the extremely radioresistant bacterium Deinococcus radiodurans. Mutat Res 435: 151–161. doi: 10.1016/S0921-8777(99)00044-0
|
| [21] |
Satoh K, Ohba H, Sghaier H, et al. (2006) Down-regulation of radioresistance by LexA2 in Deinococcus radiodurans. Microbiology 152: 3217–3226. doi: 10.1099/mic.0.29139-0
|
| [22] |
Satoh K, Tu Z, Ohba H, et al. (2009) Development of versatile shuttle vectors for Deinococcus grandis. Plasmid 62: 1–9. doi: 10.1016/j.plasmid.2009.01.005
|
 microbiol-05-02-176-s001.pdf microbiol-05-02-176-s001.pdf |
 |
| microbiol-05-02-176-s002.pdf |
|
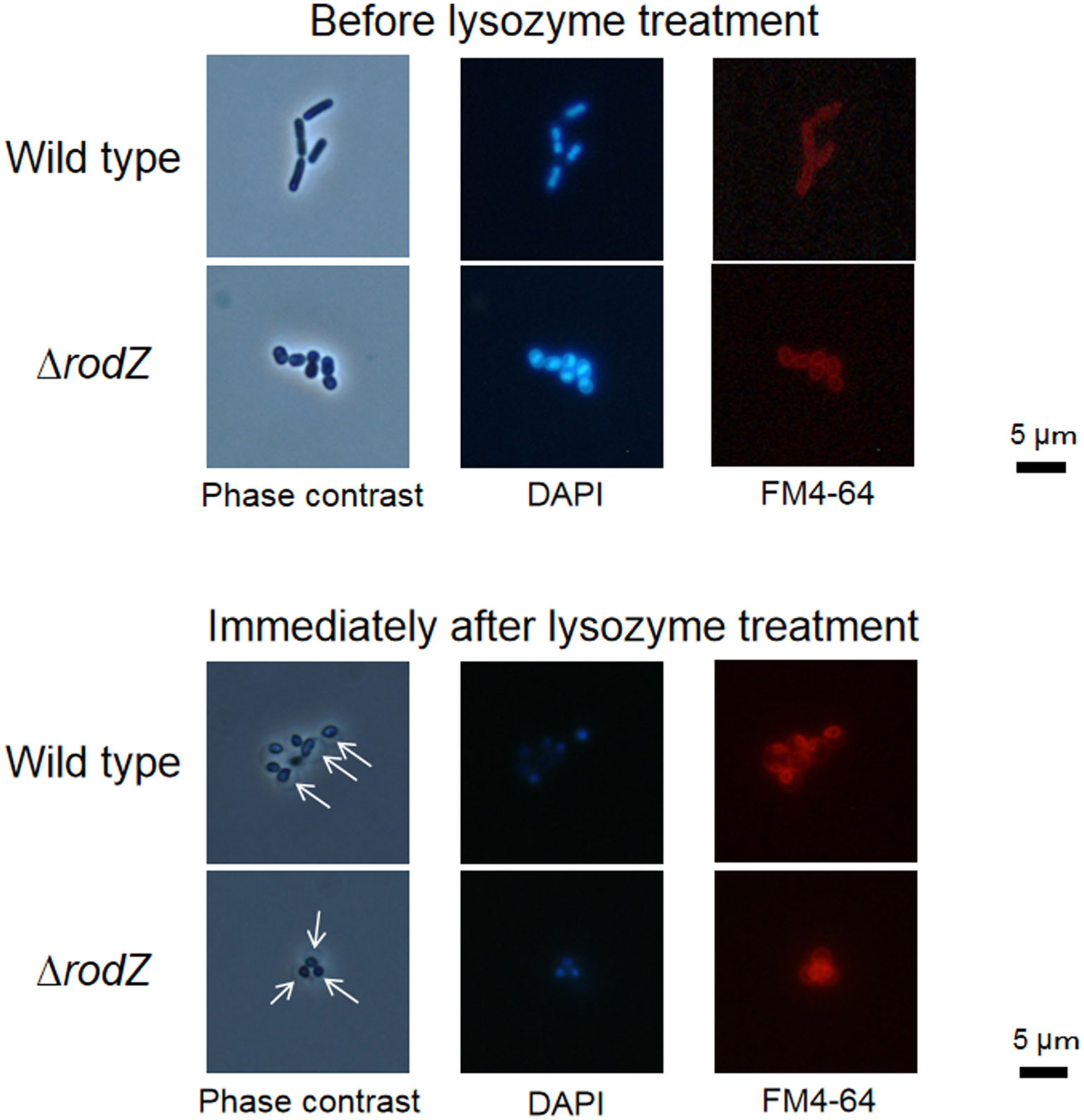
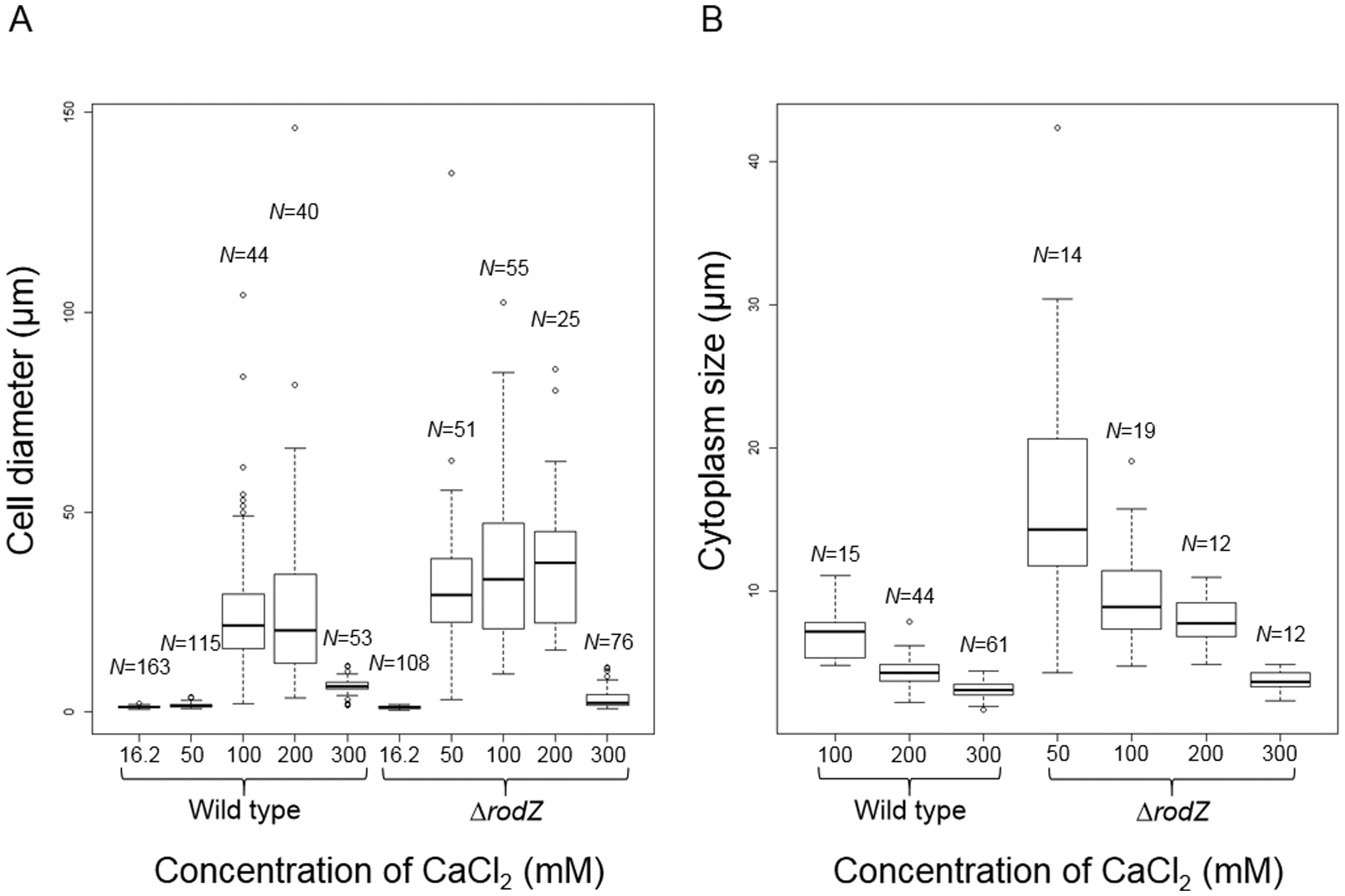
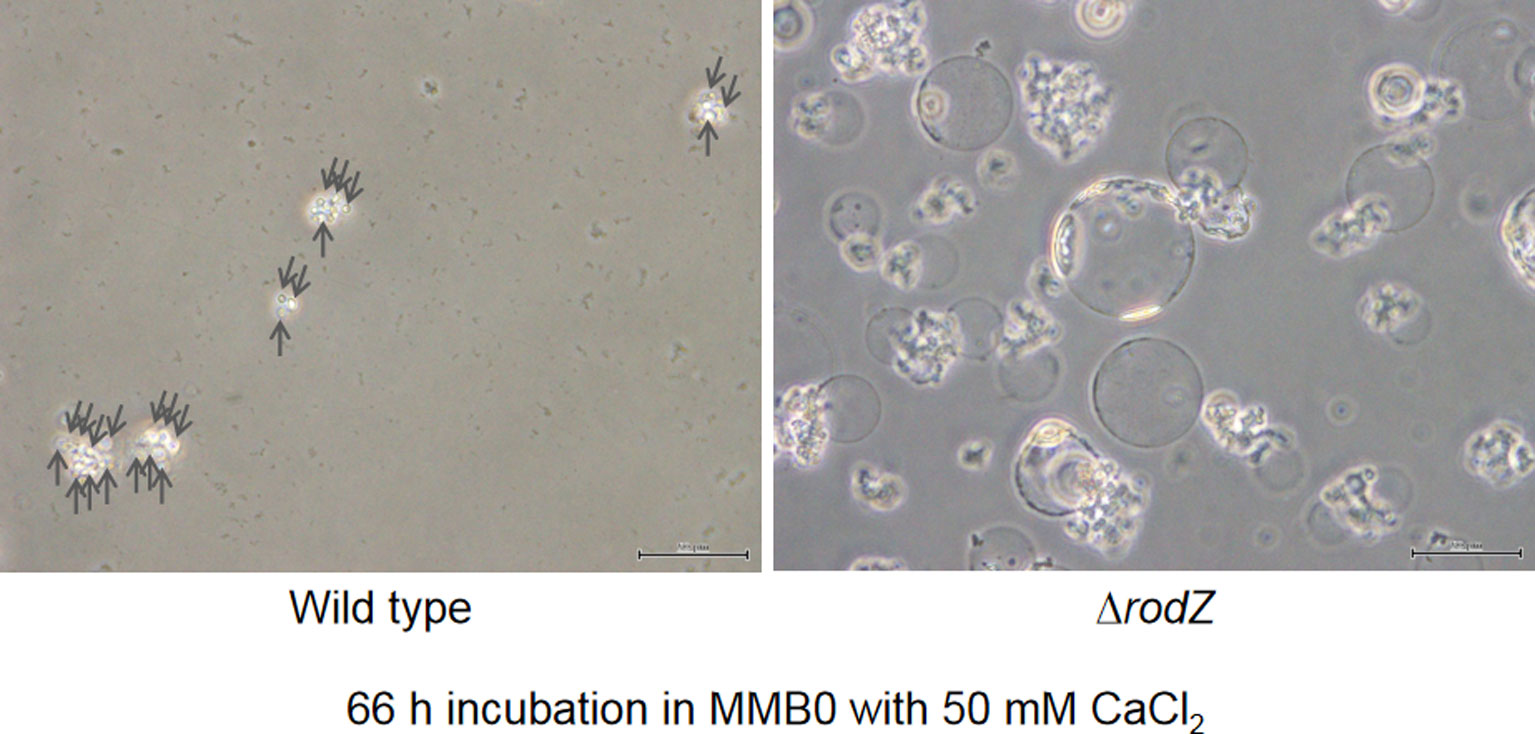
Figures(3) / Tables(1)

Yusuke Morita, Mai Okumura, Issay Narumi, Hiromi Nishida. Sensitivity of Deinococcus grandis rodZ deletion mutant to calcium ions results in enhanced spheroplast size[J]. AIMS Microbiology, 2019, 5(2): 176-185. doi: 10.3934/microbiol.2019.2.176







 DownLoad:
DownLoad: