Citation: Zoya Samoilova, Alexey Tyulenev, Nadezhda Muzyka, Galina Smirnova, Oleg Oktyabrsky. Tannic and gallic acids alter redox-parameters of the medium and modulate biofilm formation[J]. AIMS Microbiology, 2019, 5(4): 379-392. doi: 10.3934/microbiol.2019.4.379

| [1] |
Sharma G, Sharma S, Sharma P, et al. (2016) Escherichia coli biofilm: development and therapeutic strategies. J Appl Microbiol 121: 309–319. doi: 10.1111/jam.13078

|
| [2] |
Kang J, Li Q, Liu L, et al. (2018) The specific effect of gallic acid on Escherichia coli biofilm formation by regulating pgaABCD genes expression. Appl Microbiol Biotechnol 102: 1837–1846. doi: 10.1007/s00253-017-8709-3
|
| [3] |
Dong G, Liu H, Zhang X, et al. (2018) Antimicrobial and anti-biofilm activity of tannic acid against Staphylococcus aureus. Nat Prod Res 32: 2225–2228. doi: 10.1080/14786419.2017.1366485
|
| [4] |
Lee JH, Park JH, Cho HH, et al. (2013) Anti-biofilm activities of quercetin and tannic acid against Staphylococcus aureus. Biofouling 29: 491–499. doi: 10.1080/08927014.2013.788692
|
| [5] |
Borges A, Saavedra MJ, Simoes M (2012) The activity of ferulic and gallic acid on biofilm prevention and control of pathogenic bacteria. Biofouling 28: 755–767. doi: 10.1080/08927014.2012.706751
|
| [6] |
Chirwa NT, Herrington MB (2003) CsgD, a regulator of curli and cellulose synthesis, also regulates serine hydroxymethyltransferase synthesis in Escherichia coli K-12. Microbiology 149: 525–535. doi: 10.1099/mic.0.25841-0
|
| [7] |
Goulter-Thorsen RM, Taran E, Gentle IR, et al. (2011) CsgA production by Escherichia coli O157:H7 alters attachment to abiotic surfaces in some growth environments. Appl Environ Microbiol 77: 7339–7344. doi: 10.1128/AEM.00277-11
|
| [8] |
Itoh Y, Rice JD, Goller C, et al. (2008) Roles of pgaABCD genes in synthesis, modification, and export of the Escherichia coli biofilm adhesin poly-beta-1,6-N-acetyl-D-glucosamine. J Bacteriol 190: 3670–3680. doi: 10.1128/JB.01920-07
|
| [9] | Navasa N, Rodriguez-Aparicio L, Ferrero MA, et al. (2013) Polysialic and colanic acids metabolism in Escherichia coli K92 is regulated by RcsA and RcsB. Biosci Rep 33: 405–415. |
| [10] |
Jenal U, Reinders A, Lori C (2017) Cyclic di-GMP: second messenger extraordinaire. Nat Rev Microbiol 15: 271–284. doi: 10.1038/nrmicro.2016.190
|
| [11] | Herbst S, Lorkowski M, Sarenko O, et al. (2018) Transmembrane redox and proteolysis control of c-di-GMP signaling in bacterial biofilm formation. EMBO J 38: e97825. |
| [12] |
Hufnagel DA, DePas WH, Chapman M (2014) The disulfide bonding system suppresses CsgD-independent cellulose production in Escherichia coli. J Bacteriol 196: 3690–3699. doi: 10.1128/JB.02019-14
|
| [13] |
Eghbaliferiz S, Iranshahi M (2016) Prooxidant activity of polyphenols, flavonoids, anthocyanins and carotenoids: uptake review of mechanisms and catalyzing metals. Phytother Res 30: 1379–1391. doi: 10.1002/ptr.5643
|
| [14] |
Vauzour D, Rodriguez-Mateos A, Corona G, et al. (2010) Polyphenols and human health: prevention of disease and mechanisms of action. Nutrients 2: 1106–1131. doi: 10.3390/nu2111106
|
| [15] |
Ozdal T, Sela DA, Xiao J, et al. (2016) The reciprocal interactions between polyphenols and gut microbiota and effects on bioaccessibility. Nutrients 8: 78. doi: 10.3390/nu8020078
|
| [16] | Samoilova Z, Smirnova G, Oktyabrsky O (2019) The effect of catechin, tannic and gallic acids on biofilm formation in Escherichia coli depends on medium composition. Asian J Microbiol Biotechnol 4: 16–23. |
| [17] | Baba T, Ara T, Hasegawa M, et al. (2006) Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: the Keio Collection. Mol Syst Biol 2: 2006.0008. |
| [18] |
Tao K, Makino K, Yonei S, et al. (1989) Molecular cloning and nucleotide sequencing of oxyR, the positive regulatory gene of a regulon for an adaptive response to oxidative stress in Escherichia coli: homologies between OxyR protein and a family of bacterial activator proteins. Mol Gen Genet 218: 371–376. doi: 10.1007/BF00332397
|
| [19] |
Mulvey MR, Switala J, Borys A, et al. (1990) Regulation of transcription of katE and katF in Escherichia coli. J Bacteriol 172: 6713–6720. doi: 10.1128/jb.172.12.6713-6720.1990
|
| [20] | Miller JH (1972) Experiments in molecular genetics. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press. |
| [21] | Samoliova Z, Smirnova G, Oktyabrsky O (2019) Synergistic and antagonistic effects of plant extracts and antibiotics on mature Escherichia coli biofilms. Plant Cell Biol Mol Biol 20: 81–94. |
| [22] |
O'Toole GA, Kolter R (1998) Initiation of biofilm formation in Pseudomonas fluorescens WCS365 proceeds via multiple, convergent signaling pathways: A genetic analysis. Mol Microbiol 28: 449–461. doi: 10.1046/j.1365-2958.1998.00797.x
|
| [23] |
Naves P, del Prado G, Huelves L, et al. (2008) Measurement of biofilm formation by clinical isolates of Escherichia coli is method-dependent. J Appl Microbiol 105: 585–590. doi: 10.1111/j.1365-2672.2008.03791.x
|
| [24] |
Wickens HJ, Pinney RJ, Mason DJ, et al. (2000) Flow cytometric investigation of filamentation, membrane patency and membrane potential in Escherichia coli following ciprofloxacin exposure. Antimicrob Agents Chemother 44: 682–687. doi: 10.1128/AAC.44.3.682-687.2000
|
| [25] |
Smirnova GV, Muzyka NG, Ushakov VY, et al. (2015) Extracellular superoxide provokes glutathione efflux from Escherichia coli cells. Res Microbiol 166: 609–617. doi: 10.1016/j.resmic.2015.07.007
|
| [26] | Simmons L, Foti J, Cohen S, et al. (2008) The SOS regulatory network. Ecosal Plus 3 |
| [27] |
Oktyabrsky O, Vysochina G, Muzyka N, et al. (2009) Assessment of antioxidant activity of plant extracts using microbial test systems. J Appl Microbiol 106: 1175–1183. doi: 10.1111/j.1365-2672.2008.04083.x
|
| [28] |
Smirnova GV, Samoylova ZY, Muzyka NG, et al. (2009) Influence of polyphenols on Escherichia coli resistance to oxidative stress. Free Rad Biol Med 46: 759–768. doi: 10.1016/j.freeradbiomed.2008.11.017
|
| [29] |
Yen GC, Duh PD, Tsai HL (2002) Antioxidant and prooxidant effects of ascorbic acid and gallic acid. Food Chem 79: 307–313. doi: 10.1016/S0308-8146(02)00145-0
|
| [30] |
Jefferson KK (2004) What drives a bacteria to produce a biofilm? FEMS Microbiol Lett 236: 163–173. doi: 10.1111/j.1574-6968.2004.tb09643.x
|
| [31] |
Branda SS, Vik S, Friedman L, et al. (2005) Biofilms: the matrix revisited. Trends Microbiol 13: 20–26. doi: 10.1016/j.tim.2004.11.006
|
| [32] | Andrews GP, Maurelli AT (1992) mxiA of Shigella flexneri 2a, which facilitates export of invasion plasmid antigens, encodes a homologue of the low-calcium-response protein, LcrD, of Yersinia pestis. Infect Immun 60: 3287–3295. |
| [33] |
Reichhardt C, Jacobson AN, Maher MC, et al. (2015) Congo red interactions with curli-producing Escherichia coli and native curli amyloid fibers. PLoS One 10: e0140388. doi: 10.1371/journal.pone.0140388
|
| [34] |
Reichhardt C, McCrate OA, Zhou X, et al. (2016) Influence of the amyloid dye Congo red on curli, cellulose, and the extracellular matrix in Escherichia coli during growth and matrix purification. Anal Bioanal Chem 408: 7709–7717. doi: 10.1007/s00216-016-9868-2
|
| [35] | Jin ZC, Qian J (1994) Inhibitory effects of fifteen kinds of Chineese herbal drugs, vegetables and chemicals on SOS response. Zhonghua Yu Fang Yi Xue Za Zhi 28: 147–150. |
| [36] |
Khan NS, Hadi SM (1998) Structural features of taanic acid important for DNA degradation in the presence of Cu (II). Mutagenesis 13: 271–274. doi: 10.1093/mutage/13.3.271
|
| [37] |
Myszka K, Schmidt MT, Bialas W, et al. (2016) Role of gallic and p-coumaric acids in the AHL-dependent expression of flgA gene and in the process of biofilm formation in food-associated Pseudomonas fluorescens KM120. J Sci Food Agric 96: 4037–4047. doi: 10.1002/jsfa.7599
|
| [38] |
Adams JL, McLean RJ (1999) Impact of rpoS deletion on Escherichia coli biofilms. Appl Environ Microbiol 65: 4285–4287. doi: 10.1128/AEM.65.9.4285-4287.1999
|
| [39] |
Corona-Izquirdo FP, Membrillo-Hernandez J (2002) A mutation in rpoS enhances biofilm formation in Escherichia coli during exponential phase of growth. FEMS Microbiol Lett 211: 105–110. doi: 10.1111/j.1574-6968.2002.tb11210.x
|
| [40] |
Samoilova Z, Muzyka N, Lepekhina E, et al. (2014) Medicinal plant extracts can variously modify biofilm formation in Escherichia coli. Antonie van Leeuwenhoek 105: 709–722. doi: 10.1007/s10482-014-0126-3
|
| [41] |
Danese PN, Pratt LA, Kolter R (2000) Exopolysaccharide production is required for development of Escherichia coli K-12 biofilm architecture. J Bacteriol 182: 3593–3596. doi: 10.1128/JB.182.12.3593-3596.2000
|
| [42] |
Andreassen PR, Pettersen JS, Szczerba M, et al. (2018) sRNA-dependent control of curli biosynthesis in Escherichia coli: McaS directs endonucleolytic cleavage of csgD mRNA. Nucleic Acids Res 46: 6746–6760. doi: 10.1093/nar/gky479
|
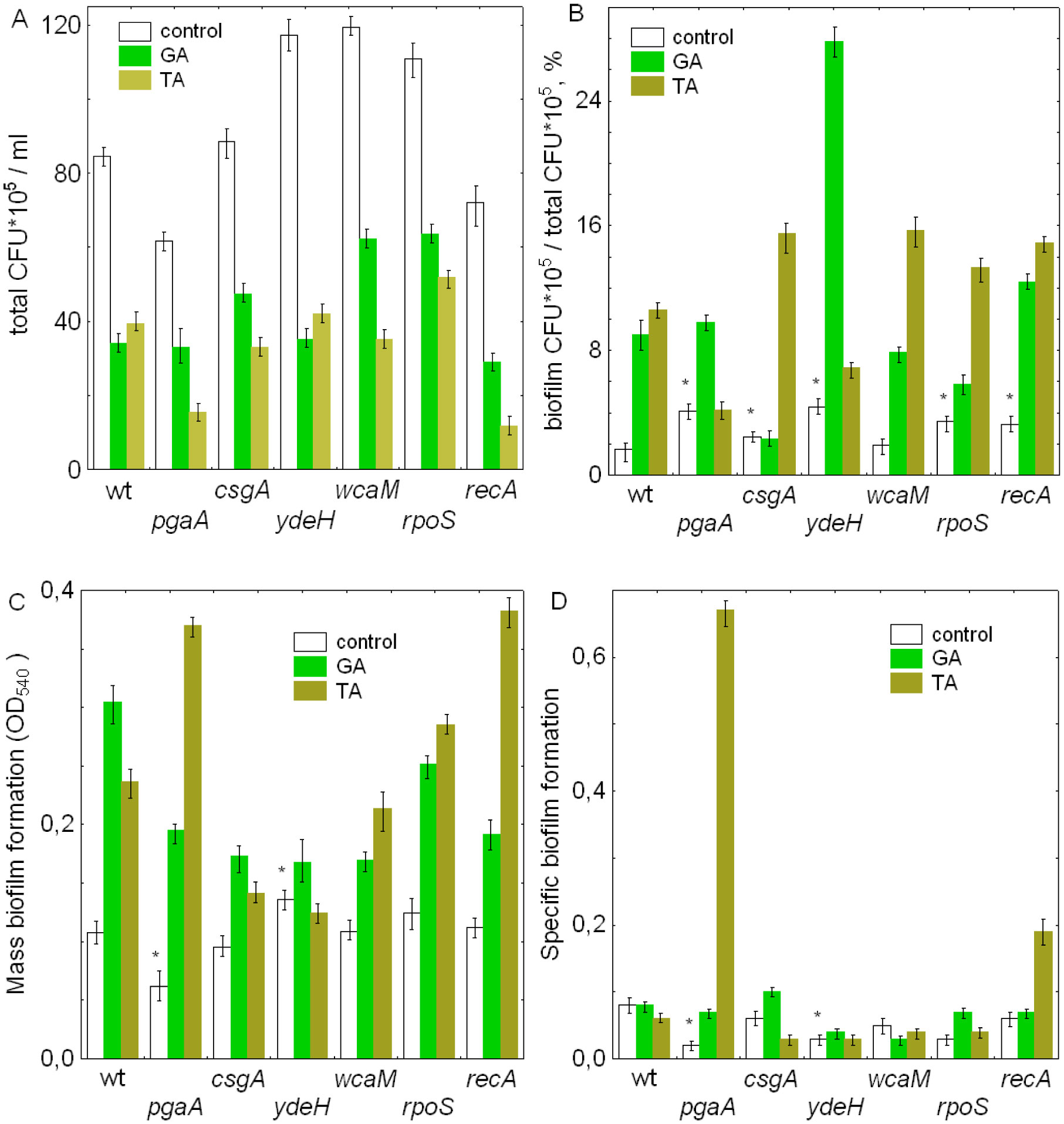
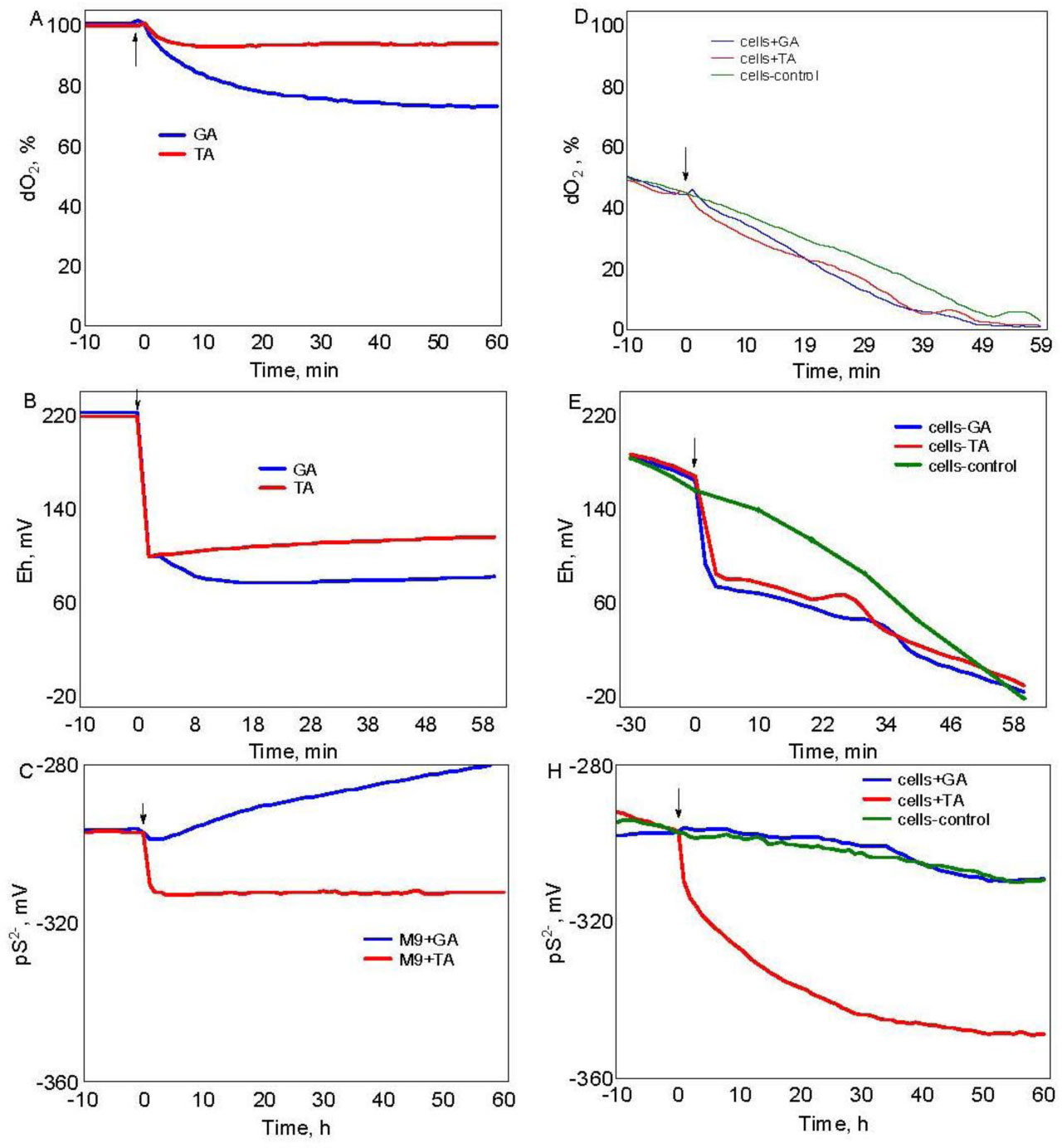
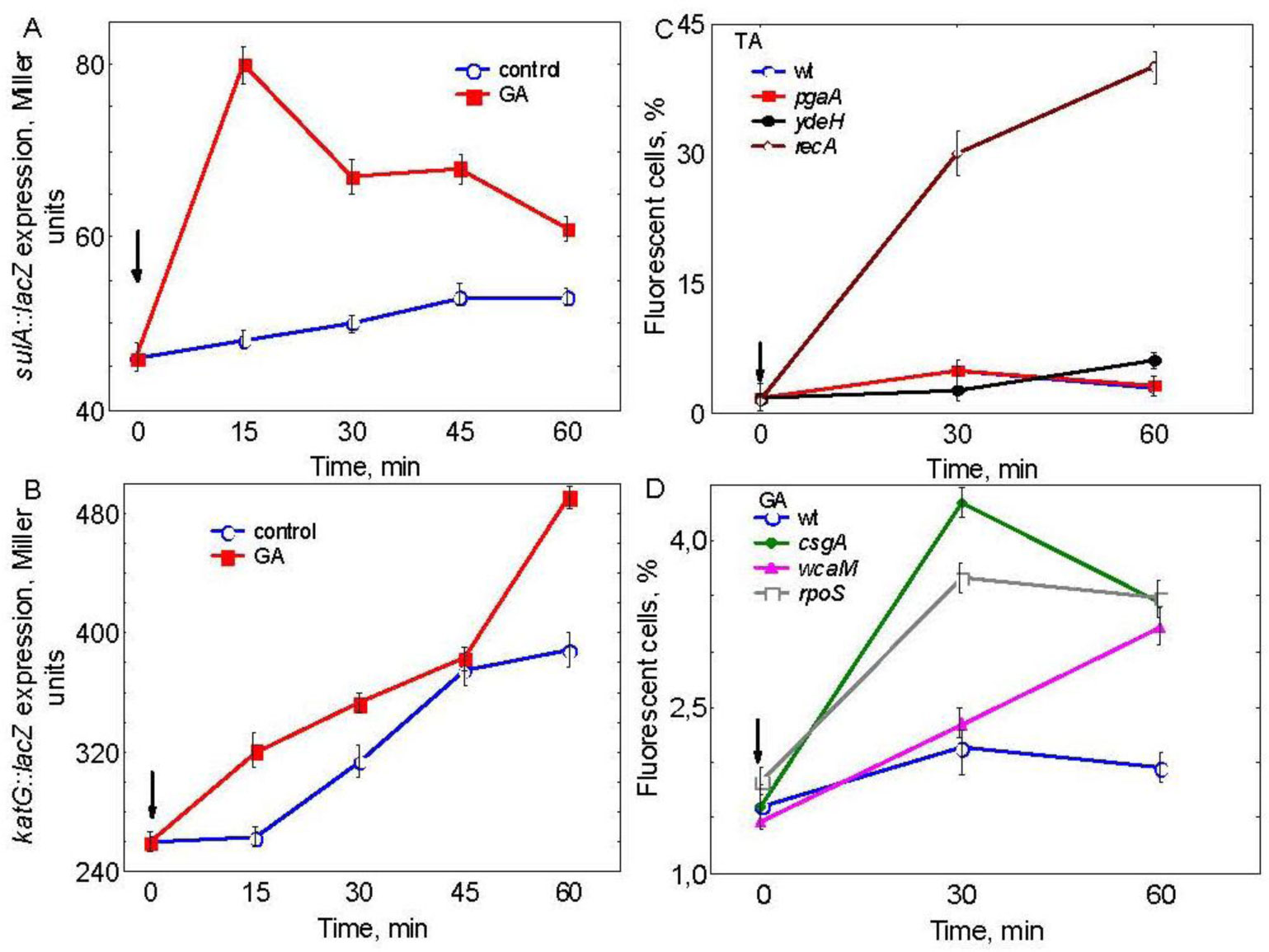
Figures(3)

Zoya Samoilova, Alexey Tyulenev, Nadezhda Muzyka, Galina Smirnova, Oleg Oktyabrsky. Tannic and gallic acids alter redox-parameters of the medium and modulate biofilm formation[J]. AIMS Microbiology, 2019, 5(4): 379-392. doi: 10.3934/microbiol.2019.4.379







 DownLoad:
DownLoad: