Citation: Tatyana V. Polyudova, Daria V. Eroshenko, Vladimir P. Korobov. Plasma, serum, albumin, and divalent metal ions inhibit the adhesion and the biofilm formation of Cutibacterium (Propionibacterium) acnes[J]. AIMS Microbiology, 2018, 4(1): 165-172. doi: 10.3934/microbiol.2018.1.165

| [1] | Scholz CF, Kilian M (2016) The natural history of cutaneous propionibacteria and reclassification of selected species within the genus Propionibacterium to the proposed novel genera Acidipropionibacterium gen. nov., Cutibacterium gen. nov. and Pseudopropionibacterium gen. nov. Int J Syst Evol Microbiol 66: 4422–4432. |
| [2] |
Christensen GJ, Brüggemann H (2014) Bacterial skin commensals and their role as host guardians. Benef Microbes 5: 201–215. doi: 10.3920/BM2012.0062

|
| [3] | Portillo ME, Corvec S, Borens O, et al. (2013) Propionibacterium acnes: An underestimated pathogen in implant-associated infections. Biomed Res Int 2013: 804391. |
| [4] |
Aubin GG, Portillo ME, Trampuz А, et al. (2014) Propionibacterium acnes, an emerging pathogen: From acne to implant-infections, from phylotype to resistance. Med Maladies Infect 44: 241–250. doi: 10.1016/j.medmal.2014.02.004
|
| [5] |
Piper KE, Jacobson MJ, Cofield RH, et al. (2009) Microbiologic diagnosis of prosthetic shoulder infection by use of implant sonication. J Clin Microbiol 47: 1878–1884. doi: 10.1128/JCM.01686-08
|
| [6] |
Coscia MF, Denys GA, Wack MF (2016) Propionibacterium acnes, coagulase-negative staphylococcus, and the "biofilm-like" intervertebral disc. Spine 41: 1860–1865. doi: 10.1097/BRS.0000000000001909
|
| [7] |
Capoor MN, Ruzicka F, Schmitz JE, et al. (2017) Propionibacterium acnes biofilm is present in intervertebral discs of patients undergoing microdiscectomy. PLoS One 12: e0174518. doi: 10.1371/journal.pone.0174518
|
| [8] | Karau MJ, Greenwoodquaintance KE, Schmidt SM, et al. (2013) Microbial biofilms and breast tissue expanders. Biomed Res Int 2013: 254940. |
| [9] |
Van VR, De RA, Verkaik NJ, et al. (2016) Prosthetic valve endocarditis due to Propionibacterium acnes. Interact Cardiov Th 23: 150–154. doi: 10.1093/icvts/ivw087
|
| [10] |
Gupta V, Yassin MH (2013) Infection and hemodialysis access: An updated review. Infect Disord Drug Targets 13: 196–205. doi: 10.2174/1871526511313030008
|
| [11] |
Holmberg A, Lood R, Mörgelin M, et al. (2009) Biofilm formation by Propionibacterium acnes is a characteristic of invasive isolates. Clin Microbiol Infect 15: 787–795. doi: 10.1111/j.1469-0691.2009.02747.x
|
| [12] | Bayston R, Ashraf W, Tucker E, et al. (2007) Biofilm formation by Propionibacterium acnes on biomaterials in vitro and in vivo: Impact on diagnosis and treatment. J Biomed Mater Res A 81: 705–709. |
| [13] | Patel JD, Ebert M, Ward R, et al. (2007) S. epidermidis biofilm formation: Effects of biomaterial surface chemistry and serum proteins. J Biomed Mater Res A 80: 742–751. |
| [14] |
Yu JL, Mânsson R, Flock JI, et al. (1997) Fibronectin binding by Propionibacterium acnes. FEMS Immunol Med Microbiol 19: 247–253. doi: 10.1111/j.1574-695X.1997.tb01094.x
|
| [15] |
Shukla SK, Rao TS (2013) Effect of calcium on Staphylococcus aureus biofilm architecture: A confocal laser scanning microscopic study. Colloid Surface B 103: 448–454. doi: 10.1016/j.colsurfb.2012.11.003
|
| [16] | Oknin H, Steinberg D, Shemesh M (2015) Magnesium ions mitigate biofilm formation of Bacillus species via downregulation of matrix genes expression. Front Microbiol 6: 907. |
| [17] |
Wu C, Labrie J, Tremblay YDN, et al. (2013) Zinc as an agent for the prevention of biofilm formation by pathogenic bacteria. J Appl Microbiol 115: 30–40. doi: 10.1111/jam.12197
|
| [18] |
El-baz R, Rizk DE, Barwa R, et al. (2017) Virulence characteristics and molecular relatedness of methicillin resistant Staphylococcus aureus harboring different staphylococcal cassette chromosome mec. Microb Pathogenesis 113: 385–395. doi: 10.1016/j.micpath.2017.11.021
|
| [19] |
Shin PK, Pawar P, Konstantopoulos K, et al. (2005) Characteristics of new Staphylococcus aureus-RBC adhesion mechanism independent of fibrinogen and IgG under hydrodynamic shear conditions. Am J Physiol-Cell Ph 289: C727–C734. doi: 10.1152/ajpcell.00034.2005
|
| [20] |
Stepanović S, Vuković D, Hola V, et al. (2007) Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. Apmis 115: 891–899. doi: 10.1111/j.1600-0463.2007.apm_630.x
|
| [21] | Rupp ME, Archer GL (1992) Hemagglutination and adherence to plastic by Staphylococcus epidermidis. Infect Immun 60: 4322–4327. |
| [22] |
She P, Chen L, Qi Y, et al. (2016) Effects of human serum and apo-Transferrin on Staphylococcus epidermidis RP62A biofilm formation. MicrobiologyOpen 5: 957–966. doi: 10.1002/mbo3.379
|
| [23] |
Bae JY, Park SN (2016) Evaluation of anti-microbial activities of ZnO, citric acid and a mixture of both against Propionibacterium acnes. Int J Cosmet Sci 38: 550–557. doi: 10.1111/ics.12318
|
| [24] |
Geesey GG, Wigglesworth-Cooksey B, Cooksey KE (2000) Influence of calcium and other cations on surface adhesion of bacteria and diatoms: A review. Biofouling 15: 195–205. doi: 10.1080/08927010009386310
|
| [25] |
Formosa-Dague C, Speziale P, Foster TJ, et al. (2016) Zinc-dependent mechanical properties of Staphylococcus aureus biofilm-forming surface protein SasG. Proc Natl Acad Sci USA 113: 410–415. doi: 10.1073/pnas.1519265113
|
| [26] |
Ozuguz P, Kacar SD, Ekiz O, et al. (2014) Evaluation of serum vitamins A and E and zinc levels according to the severity of acne vulgaris. J Toxicol-Cutan Ocu 33: 99–102. doi: 10.3109/15569527.2013.808656
|
| [27] |
Prasad AS, Mantzoros CS, Beck FW, et al. (1996) Zinc status and serum testosterone levels of healthy adults. Nutrition 12: 344–348. doi: 10.1016/S0899-9007(96)80058-X
|
| [28] |
Aubin GG, Baud'huin M, Lavigne JP, et al. (2017) Interaction of Cutibacterium (formerly Propionibacterium) acnes with bone cells: A step toward understanding bone and joint infection development. Sci Rep 7: 42918. doi: 10.1038/srep42918
|
| [29] |
Cousin F, Deutsch SM, Perez CA, et al. (2012) Interactions between probiotic dairy propionibacteria and the intestinal epithelium. Curr Immunol Rev 8: 216–226. doi: 10.2174/157339512800671976
|
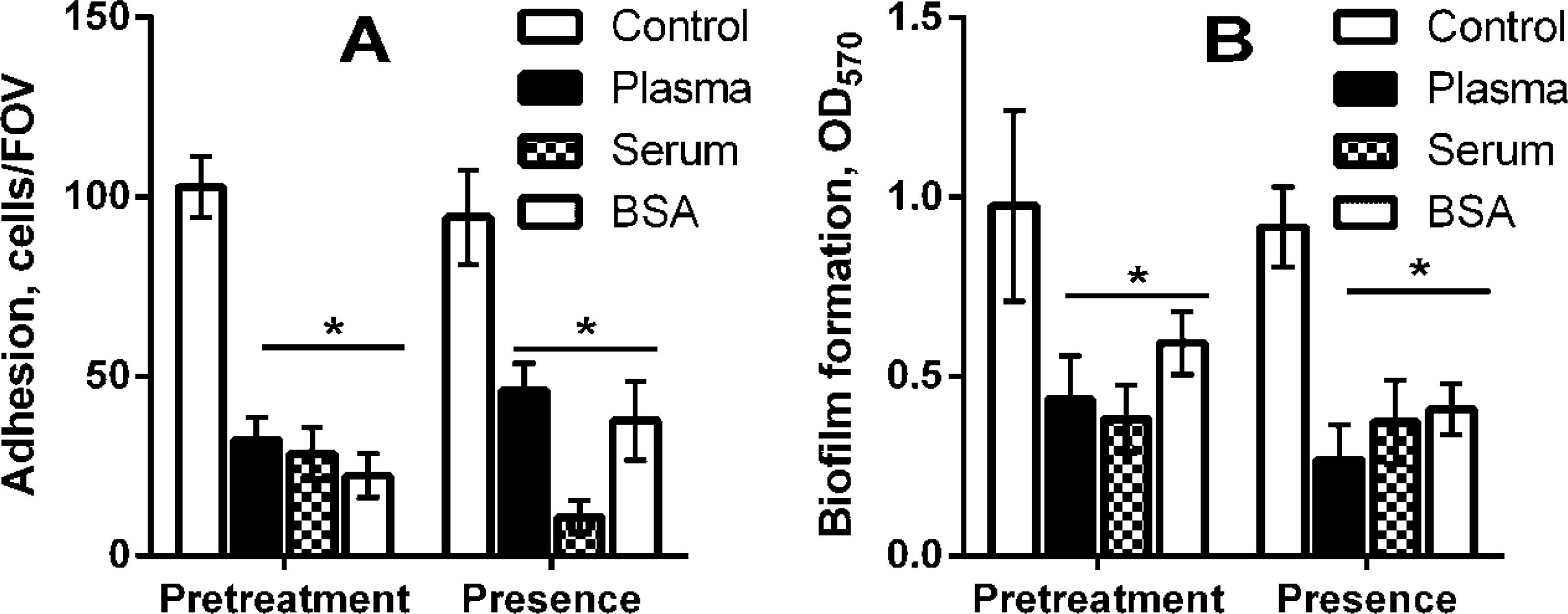
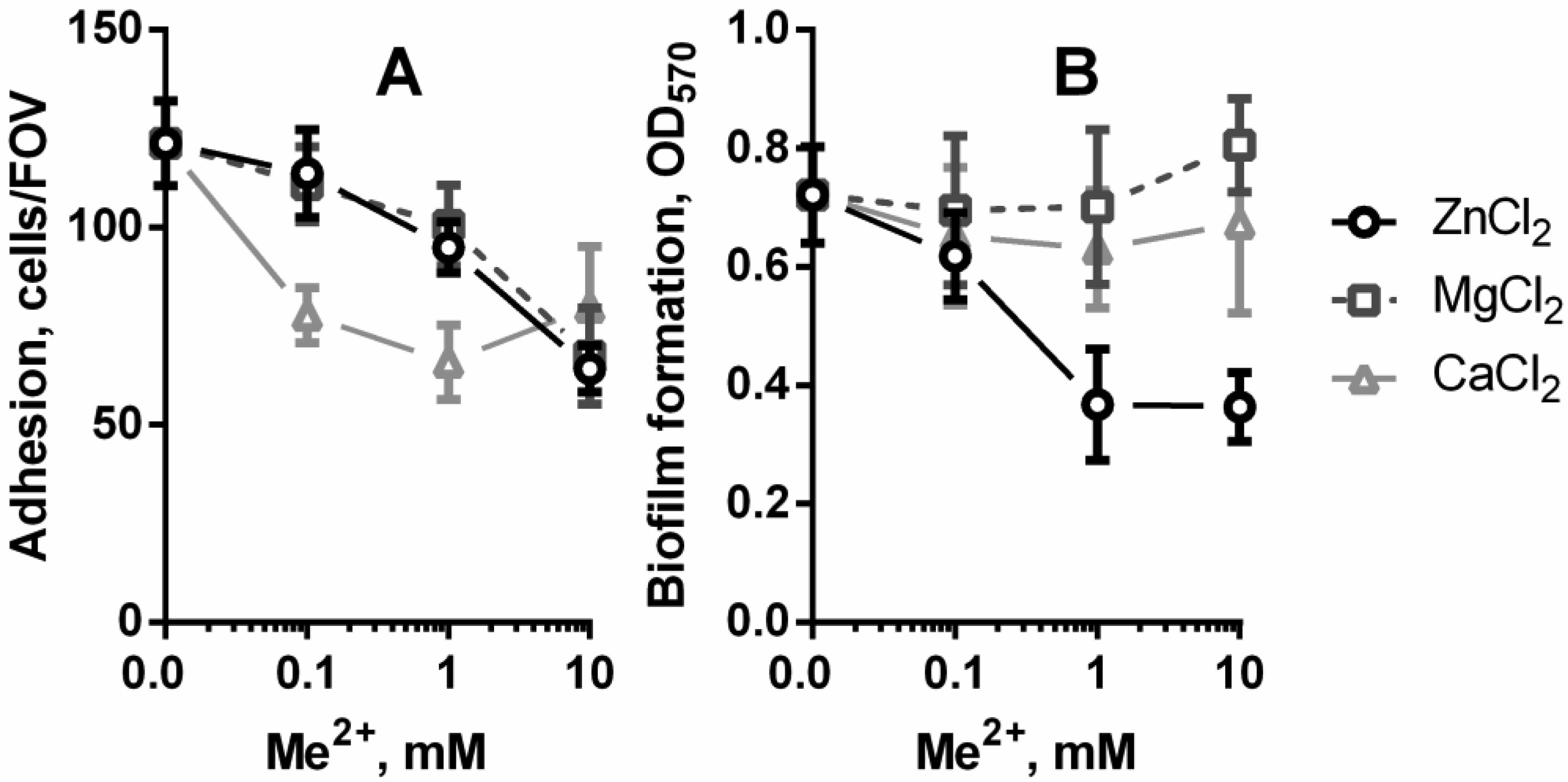
Figures(3)

Tatyana V. Polyudova, Daria V. Eroshenko, Vladimir P. Korobov. Plasma, serum, albumin, and divalent metal ions inhibit the adhesion and the biofilm formation of Cutibacterium (Propionibacterium) acnes[J]. AIMS Microbiology, 2018, 4(1): 165-172. doi: 10.3934/microbiol.2018.1.165







 DownLoad:
DownLoad: