Citation: Laurent Coquet, Antoine Obry, Nabil Borghol, Julie Hardouin, Laurence Mora, Ali Othmane, Thierry Jouenne. Impact of chlorhexidine digluconate and temperature on curli production in Escherichia coli—consequence on its adhesion ability[J]. AIMS Microbiology, 2017, 3(4): 915-937. doi: 10.3934/microbiol.2017.4.915

| [1] | Pinzauti S (1983) Chlorhexidine in dentistry, preservation of contact lens and skin disinfection. Farmaco Prat 38: 21–42. |
| [2] |
Lee I, Agarwal RK, Lee BY, et al. (2010) Systematic review and cost analysis comparing use of chlorhexidine with use of iodine for preoperative skin antisepsis to prevent surgical site infection. Infect Cont Hosp Ep 31: 1219–1229. doi: 10.1086/657134

|
| [3] |
Garrett TR, Bhakoo M, Zhang Z (2008) Bacterial adhesion and biofilms on surfaces. Prog Nat Sci 18: 1049–1056. doi: 10.1016/j.pnsc.2008.04.001
|
| [4] |
Goulter RM, Gentle IR, Dykes GA (2009) Issues in determining factors influencing bacterial attachment: a review using the attachment of Escherichia coli to abiotic surfaces as an example. Lett Appl Microbiol 49: 1–7. doi: 10.1111/j.1472-765X.2009.02591.x
|
| [5] |
Zhou Y, Smith DR, Hufnagel DA, et al. (2013) Experimental manipulation of the microbial functional amyloid called curli. Methods Mol Biol 966: 53–75. doi: 10.1007/978-1-62703-245-2_4
|
| [6] |
Zogaj X, Nimtz M, Rohde M, et al. (2001) The multicellular morphotypes of Salmonella typhimurium and Escherichia coli produce cellulose as the second component of the extracellular matrix. Mol Microbiol 39: 1452–1463. doi: 10.1046/j.1365-2958.2001.02337.x
|
| [7] |
Barnhart MM, Chapman MR (2006) Curli biogenesis and function. Annu Rev Microbiol 60: 131–147. doi: 10.1146/annurev.micro.60.080805.142106
|
| [8] |
Jubelin G, Vianney A, Beloin C, et al. (2005) CpxR/OmpR interplay regulates curli gene expression in response to osmolarity in Escherichia coli. J Bacteriol 187: 2038–2049. doi: 10.1128/JB.187.6.2038-2049.2005
|
| [9] | Olsén A, Arnqvist A, Hammar M, et al. (1993) Environmental regulation of curli production in Escherichia coli. Infect Agents Dis 2: 272–274. |
| [10] | Hufnagel DA, Depas WH, Chapman MR (2015) The biology of the Escherichia coli extracellular matrix. Microbiol Spectrum 3. |
| [11] | Evans ML, Chapman MR (2014) Curli biogenesis: order out of disorder. BBA-Mol Cell Res 1843: 1551–1558. |
| [12] |
Prigent-Combaret C, Brombacher E, Vidal O, et al. (2001) Complex regulatory network controls initial adhesion and biofilm formation in Escherichia coli via regulation of the csgD gene. J Bacteriol 183: 7213–7223. doi: 10.1128/JB.183.24.7213-7223.2001
|
| [13] |
Dorel C, Lejeune P, Rodrigue A (2006) The Cpx system of Escherichia coli, a strategic signaling pathway for confronting adverse conditions and for settling biofilm communities? Res Microbiol 157: 306–314. doi: 10.1016/j.resmic.2005.12.003
|
| [14] | Vidal O, Longin R, Prigent-Combaret C, et al. (1998) Isolation of an Escherichia coli K-12 mutant strain able to form biofilms on inert surfaces: involvement of a new ompR allele that increases curli expression. J Bacteriol 180: 2442–2449. |
| [15] |
Uhlich GA, Cooke PH, Solomon EB (2006) Analyses of the red-dry-rough phenotype of an Escherichia coli O157:H7 strain and its role in biofilm formation and resistance to antibacterial agents. Appl Environ Microb 72: 2564–2572. doi: 10.1128/AEM.72.4.2564-2572.2006
|
| [16] |
Saldaña Z, Xicohtencatl-Cortes J, Avelino F, et al. (2009) Synergistic role of curli and cellulose in cell adherence and biofilm formation of attaching and effacing Escherichia coli and identification of Fis as a negative regulator of curli. Environ Microbiol 11: 992–1006. doi: 10.1111/j.1462-2920.2008.01824.x
|
| [17] | Macé C, Seyer D, Chemani C, et al. (2008) Identification of biofilm-associated cluster (bac) in Pseudomonas aeruginosa involved in biofilm formation and virulence. PLoS One 3: e0003897. |
| [18] |
Coquet L, Cosette P, Quillet L, et al. (2002) Occurrence and phenotypic characterization of Yersinia ruckeri strains with biofilm-forming capacity in a rainbow trout farm. Appl Environ Microb 68: 470–475. doi: 10.1128/AEM.68.2.470-475.2002
|
| [19] |
Obry A, Lequerré T, Hardouin J, et al. (2014) Identification of S100A9 as biomarker of responsiveness to the methotrexate/etanercept combination in rheumatoid arthritis using a proteomic approach. PLoS One 9: e115800. doi: 10.1371/journal.pone.0115800
|
| [20] |
Amorim CVG, Aun CE, Mayer MPA (2004) Susceptibility of some oral microorganisms to chlorhexidine and paramonochlorophenol. Braz Oral Res 18: 242–246. doi: 10.1590/S1806-83242004000300012
|
| [21] |
Shivaji S, Prakash JSS (2010) How do bacteria sense and respond to low temperature? Arch Microbiol 192: 85–95. doi: 10.1007/s00203-009-0539-y
|
| [22] |
Wojnicz D, Tichaczek-Goska D (2013) Effect of sub-minimum inhibitory concentrations of ciprofloxacin, amikacin and colistin on biofilm formation and virulence factors of Escherichia coli planktonic and biofilm forms isolated from human urine. Braz J Microbiol 44: 259–265. doi: 10.1590/S1517-83822013000100037
|
| [23] |
Kikuchi T, Mizunoe Y, Takade A, et al. (2005) Curli fibers are required for development of biofilm architecture in Escherichia coli K-12 and enhance bacterial adherence to human uroepithelial cells. Microbiol Immunol 49: 875–884. doi: 10.1111/j.1348-0421.2005.tb03678.x
|
| [24] |
Ryu JH, Kim H, Frank JF, et al. (2004) Attachment and biofilm formation on stainless steel by Escherichia coli O157:H7 as affected by curli production. Lett Appl Microbiol 39: 359–362. doi: 10.1111/j.1472-765X.2004.01591.x
|
| [25] |
Kaplan JB (2011) Antibiotic-induced biofilm formation. Int J Artif Organs 34: 737–751. doi: 10.5301/ijao.5000027
|
| [26] |
Houari A, Martino PD (2007) Effect of chlorhexidine and benzalkonium chloride on bacterial biofilm formation. Lett Appl Microbiol 45: 652–656. doi: 10.1111/j.1472-765X.2007.02249.x
|
| [27] |
White-Ziegler CA, Um S, Pérez NM, et al. (2008) Low temperature (23 degrees C) increases expression of biofilm-, cold-shock- and RpoS-dependent genes in Escherichia coli K-12. Microbiology 154: 148–166. doi: 10.1099/mic.0.2007/012021-0
|
| [28] |
Yoon SH, Han MJ, Lee SY, et al. (2003) Combined transcriptome and proteome analysis of Escherichia coli during high cell density culture. Biotechnol Bioeng 81: 753–767. doi: 10.1002/bit.10626
|
| [29] |
Cheung HY, Wong MMK, Cheung SH, et al. (2012) Differential actions of chlorhexidine on the cell wall of Bacillus subtilis and Escherichia coli. PLoS One 7: e36659. doi: 10.1371/journal.pone.0036659
|
| [30] | Condell O, Power KA, Händler K, et al. (2014) Comparative analysis of Salmonella susceptibility and tolerance to the biocide chlorhexidine identifies a complex cellular defense network. Front Microbiol 5: 373. |
| [31] | Goemans C, Denoncin K, Collet JF (2014) Folding mechanisms of periplasmic proteins. BBA-Mol Cell Res 1843: 1517–1528. |
| [32] |
Vertommen D, Ruiz N, Leverrier P, et al. (2009) Characterization of the role of the Escherichia coli periplasmic chaperone SurA using differential proteomics. Proteomics 9: 2432–2443. doi: 10.1002/pmic.200800794
|
| [33] | Zhang D, Sweredoski MJ, Graham RLJ, et al. (2012) Novel proteomic tools reveal essential roles of SRP and importance of proper membrane protein biogenesis. Mol Cell Proteomics 11: M111.011585. |
| [34] |
Price NL, Raivio TL (2009) Characterization of the Cpx Regulon in Escherichia coli strain MC4100. J Bacteriol 191: 1798–1815. doi: 10.1128/JB.00798-08
|
| [35] |
Surmann K, Ćudić E, Hammer E, et al. (2016) Molecular and proteome analyses highlight the importance of the Cpx envelope stress system for acid stress and cell wall stability in Escherichia coli. MicrobiologyOpen 5: 582–596. doi: 10.1002/mbo3.353
|
| [36] |
Carter MQ, Brandl MT, Louie JW, et al. (2011) Distinct acid resistance and survival fitness displayed by Curli variants of enterohemorrhagic Escherichia coli O157:H7. Appl Environ Microb 77: 3685–3695. doi: 10.1128/AEM.02315-10
|
| [37] |
Alexander DM, John ACS (1994) Characterization of the carbon starvation-inducible and stationary phase-inducible gene slp encoding an outer membrane lipoprotein in Escherichia coli. Mol Microbiol 11: 1059–1071. doi: 10.1111/j.1365-2958.1994.tb00383.x
|
| [38] |
Mates AK, Sayed AK, Foster JW (2007) Products of the Escherichia coli acid fitness island attenuate metabolite stress at extremely low pH and mediate a cell density-dependent acid resistance. J Bacteriol 189: 2759–2768. doi: 10.1128/JB.01490-06
|
| [39] |
Shimizu K (2013) Regulation systems of bacteria such as Escherichia coli in response to nutrient limitation and environmental stresses. Metabolites 4: 1–35. doi: 10.3390/metabo4010001
|
| [40] |
Conter A, Menchon C, Gutierrez C (1997) Role of DNA supercoiling and rpoS sigma factor in the osmotic and growth phase-dependent induction of the gene osmE of Escherichia coli K12. J Mol Biol 273: 75–83. doi: 10.1006/jmbi.1997.1308
|
| [41] |
Bäumler AJ, Hantke K (1992) A lipoprotein of Yersinia enterocolitica facilitates ferrioxamine uptake in Escherichia coli. J Bacteriol 174: 1029–1035. doi: 10.1128/jb.174.3.1029-1035.1992
|
| [42] |
Plesa M, Hernalsteens JP, Vandenbussche G, et al. (2006) The SlyB outer membrane lipoprotein of Burkholderia multivorans contributes to membrane integrity. Res Microbiol 157: 582–592. doi: 10.1016/j.resmic.2005.11.015
|
| [43] |
Murata M, Noor R, Nagamitsu H, et al. (2012) Novel pathway directed by σ E to cause cell lysis in Escherichia coli. Genes Cells 17: 234–247. doi: 10.1111/j.1365-2443.2012.01585.x
|
| [44] | Rahfeld JU, Rücknagel KP, Stoller G, et al. (1996) Isolation and amino acid sequence of a new 22-kDa FKBP-like peptidyl-prolyl cis/trans-isomerase of Escherichia coli. Similarity to Mip-like proteins of pathogenic bacteria. J Biol Chem 271: 22130–22138. |
| [45] |
Ishihama A (2010) Prokaryotic genome regulation: multifactor promoters, multitarget regulators and hierarchic networks. FEMS Microbiol Rev 34: 628–645. doi: 10.1111/j.1574-6976.2010.00227.x
|
| [46] |
Rodgers N, Murdaugh A (2016) Chlorhexidine-induced elastic and adhesive changes of Escherichia coli cells within a biofilm. Biointerphases 11: 031011. doi: 10.1116/1.4962265
|
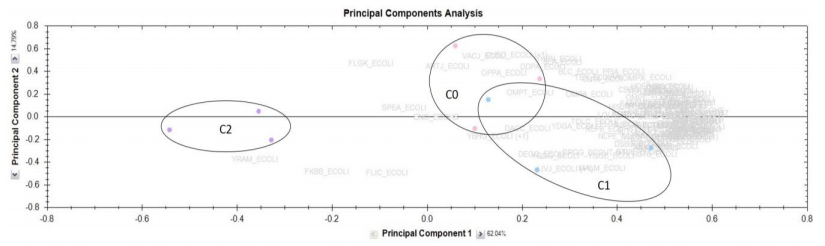
Figures(5) / Tables(2)

Laurent Coquet, Antoine Obry, Nabil Borghol, Julie Hardouin, Laurence Mora, Ali Othmane, Thierry Jouenne. Impact of chlorhexidine digluconate and temperature on curli production in Escherichia coli—consequence on its adhesion ability[J]. AIMS Microbiology, 2017, 3(4): 915-937. doi: 10.3934/microbiol.2017.4.915







 DownLoad:
DownLoad: