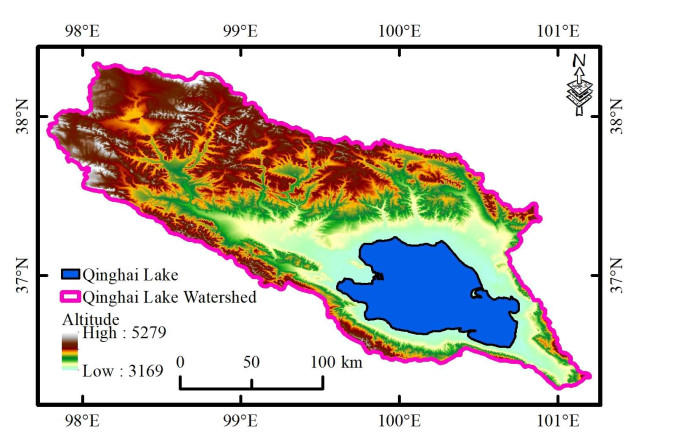
The global climate has undergone great changes in recent decades, which has a significant impact on the vegetation system, especially in arid and semi-arid areas. Based on a dynamic model, this paper studied the response of vegetation pattern to climate change in Qinghai Lake, a typical semi-arid region. The conditions for Turing instability of the equilibrium were obtained by mathematical analysis. The numerical experiments showed the influence of different climitic factors (carbon dioxide concentrations [$ CO_2 $], temperature and precipitation) on vegetation pattern. The results showed that the robustness of the vegetation system was enhanced as precipitation or [$ CO_2 $] increased. Furthermore, we presented evolution of vegetation system under different climate scenarios to forecast the future growth of vegetation. We compared the various climate scenarios with representative concentration pathways (RCP2.6, RCP4.5, RCP8.5). The results revealed that RCP2.6 scenario was a desired climate scenario for Qinghai Lake. Our study also highlighted the measures to avoid desertification by the method of optimal control. We expect that this study will provide theoretical basis for vegetation protection.
Citation: Juan Liang, Huilian Ma, Huanqing Yang, Zunguang Guo. Response of vegetation pattern to climate change based on dynamical model: Case of Qinghai Lake, China[J]. AIMS Mathematics, 2024, 9(1): 2500-2517. doi: 10.3934/math.2024123
The global climate has undergone great changes in recent decades, which has a significant impact on the vegetation system, especially in arid and semi-arid areas. Based on a dynamic model, this paper studied the response of vegetation pattern to climate change in Qinghai Lake, a typical semi-arid region. The conditions for Turing instability of the equilibrium were obtained by mathematical analysis. The numerical experiments showed the influence of different climitic factors (carbon dioxide concentrations [$ CO_2 $], temperature and precipitation) on vegetation pattern. The results showed that the robustness of the vegetation system was enhanced as precipitation or [$ CO_2 $] increased. Furthermore, we presented evolution of vegetation system under different climate scenarios to forecast the future growth of vegetation. We compared the various climate scenarios with representative concentration pathways (RCP2.6, RCP4.5, RCP8.5). The results revealed that RCP2.6 scenario was a desired climate scenario for Qinghai Lake. Our study also highlighted the measures to avoid desertification by the method of optimal control. We expect that this study will provide theoretical basis for vegetation protection.

| [1] |
Z. Wu, N. E. Huang, J. M. Wallace, B. V. Smoliak, X. Chen, On the time-varying trend in global-mean surface temperature, Clim. Dynam., 37 (2011), 759–773. https://doi.org/10.1007/s00382-011-1128-8 doi: 10.1007/s00382-011-1128-8

|
| [2] |
J. Huang, X. Guan, F. Ji, Enhanced cold-season warming in semi-arid regions, Atmos. Chem. Phys., 12 (2012), 5391–5398. https://doi.org/10.5194/acp-12-5391-2012 doi: 10.5194/acp-12-5391-2012
|
| [3] |
A. Dai, Drought under global warming: A review, WIREs. Clim. Change, 2 (2011), 45–65. https://doi.org/10.1002/wcc.81 doi: 10.1002/wcc.81
|
| [4] | J. Hansen, R. Ruedy, M. Sato, K. Lo, Global surface temperature change, Rev. Geophys., 48 (2010). https://doi.org/10.1029/2010RG000345 |
| [5] |
Z. Li, J. Gao, L. Wen, C. Zou, C. Feng, D. Li, et al., Dynamics of soil respiration in alpine wetland meadows exposed to different levels of degradation in the Qinghai-Tibet Plateau, China, Sci. Rep., 9 (2019), 7469. https://doi.org/10.1038/s41598-019-43904-1 doi: 10.1038/s41598-019-43904-1
|
| [6] |
J. Gao, H. Ouyang, G. Lei, X. Xu, M. Zhang, Effects of temperature, soil moisture, soil type and their interactions on soil carbon mineralization in Zoigo alpine wetland, Qinghai-Tibet Plateau, Chinese Geogr. Sci., 21 (2011), 27–35. https://doi.org/10.1007/s11769-011-0439-3 doi: 10.1007/s11769-011-0439-3
|
| [7] |
C. Mu, T. Zhang, Q. Zhao, H. Su, S. Wang, B. Cao, et al., Permafrost affects carbon exchange and its response to experimental warming on the northern Qinghai-Tibetan Plateau, Agr. Forest Meteorol., 247 (2017), 252–259. https://doi.org/10.1016/j.agrformet.2017.08.009 doi: 10.1016/j.agrformet.2017.08.009
|
| [8] |
F. Peng, Q. G. You, M. H. Xu, X. H. Zhou, T. Wang, G. Guo, et al., Effects of experimental warming on soil respiration and its components in an alpine meadow in the permafrost region of the Qinghai-Tibet Plateau, Eur. J. Soil Sci., 66 (2015), 145–154. https://doi.org/10.1111/ejss.12187 doi: 10.1111/ejss.12187
|
| [9] |
Z. X. Xu, T. L. Gong, J. Y. Li, Decadal trend of climate in the Tibetan Plateaui-regional temperature and precipitation, Hydrol. Process., 22 (2008), 3056–3065. https://doi.org/10.1002/hyp.6892 doi: 10.1002/hyp.6892
|
| [10] |
G. Zhang, T. Yao, H. Xie, K. Yang, L. Zhu, C. K. Shum, et al., Response of Tibetan Plateau lakes to climate change: Trends, patterns, and mechanisms, Earth Sci. Rev., 208 (2020), 103269. https://doi.org/10.1016/j.earscirev.2020.103269 doi: 10.1016/j.earscirev.2020.103269
|
| [11] |
J. Wang, Q. Wu, Impact of experimental warming on soil temperature and moisture of the shallow active layer of wet meadows on the Qinghai-Tibet Plateau, Cold Reg. Sci. Technol., 90-91 (2013), 1–8. https://doi.org/10.1016/j.coldregions.2013.03.005 doi: 10.1016/j.coldregions.2013.03.005
|
| [12] |
W. Wan, L. Zhao, H. Xie, B. Liu, H. Li, Y. Cui, et al., Lake surface water temperature change over the Tibetan plateau from 2001 to 2015: A sensitive indicator of the warming climate, Geophys. Res. Lett., 45 (2018), 11177–11186. https://doi.org/10.1029/2018GL078601 doi: 10.1029/2018GL078601
|
| [13] |
G. Wang, Y. Li, Q. Wu, Y. Wang, Impacts of permafrost changes on alpine ecosystem in Qinghai-Tibet Plateau, Sci. China Ser. D, 49 (2006), 1156–1169. https://doi.org/10.1007/s11430-006-1156-0 doi: 10.1007/s11430-006-1156-0
|
| [14] |
X. Wu, L. Zhao, M. Chen, H. Fang, G. Yue, J. Chen, et al., Soil organic carbon and its relationship to vegetation communities and soil properties in permafrost areas of the Central Western Qinghai-Tibet Plateau, China, Permafrost Periglac. Process., 23 (2012), 162–169. https://doi.org/10.1002/ppp.1740 doi: 10.1002/ppp.1740
|
| [15] |
X. Wu, H. Fang, Y. Zhao, J. M. Smoak, W. Li, W. Shi, et al., A conceptual model of the controlling factors of soil organic carbon and nitrogen densities in a permafrost-affected region on the eastern Qinghai-Tibetan Plateau, J. Geophys. Rese. Biogeo., 122 (2017), 1705–1717. https://doi.org/10.1002/2016JG003641 doi: 10.1002/2016JG003641
|
| [16] | S. Cao, G. Cao, Q. Feng, G. Han, Y. Lin, J. Yuan, et al., Alpine wetland ecosystem carbon sink and its controls at the Qinghai Lake, Environ. Earth Sci. 76 (2017), 210. https://doi.org/10.1007/s12665-017-6529-5 |
| [17] |
X. Li, W. Liu, L. Xu, Carbon isotopes in surface-sediment carbonates of modern Lake Qinghai (Qinghai-Tibet Plateau): Implications for lake evolution in arid areas, Chem. Geol., 300 (2012), 88–96. https://doi.org/10.1016/j.chemgeo.2012.01.010 doi: 10.1016/j.chemgeo.2012.01.010
|
| [18] |
H. Ao, C. Wu, X. Xiong, L. Jing, X. Huang, K. Zhang, et al., Water and sediment quality in Qinghai Lake, China: A revisit after half a century, Environ. Monit. Assess., 186 (2014), 2121–2133. https://doi.org/10.1007/s10661-013-3522-7 doi: 10.1007/s10661-013-3522-7
|
| [19] |
L. Tang, X. Duan, F. Kong, F. Zhang, Y. Zheng, Z. Li, et al., Influences of climate change on area variation of Qinghai Lake on Qinghai-Tibetan Plateau since 1980s, Sci. Rep., 8 (2018), 7331. https://doi.org/10.1038/s41598-018-25683-3 doi: 10.1038/s41598-018-25683-3
|
| [20] |
B. Chang, K. N. He, R. J. Li, Z. P. Sheng, H. Wang, Linkage of climatic factors and human activities with water level fluctuations in Qinghai Lake in the northeastern Tibetan Plateau, China, Water, 9 (2017), 552. https://doi.org/10.3390/w9070552 doi: 10.3390/w9070552
|
| [21] |
C. Fan, C. Song, W. Li, K. Liu, J. Cheng, C. Fu, et al., What drives the rapid water-level recovery of the largest lake (Qinghai Lake) of China over the past half century?, J. Hydrol., 593 (2021), 125921. https://doi.org/10.1016/j.jhydrol.2020.125921 doi: 10.1016/j.jhydrol.2020.125921
|
| [22] | H. Dong, Y. Song, M. Zhang, Hydrological trend of Qinghai Lake over the last 60 years: Driven by climate variations or human activities? J. Water Climate Change, 10 (2019), 524–534. https://doi.org/10.2166/wcc.2018.033 |
| [23] |
W. Zhang, S. Wang, B. Zhang, F. Zhang, Q. Shen, Y. Wu, et al., Analysis of the water color transitional change in Qinghai Lake during the past 35 years observed from Landsat and MODIS, J. Hydrol. Reg. Stud., 42 (2022), 101154. https://doi.org/10.1016/j.ejrh.2022.101154 doi: 10.1016/j.ejrh.2022.101154
|
| [24] |
T. Che, X. Li, R. Jin, Monitoring the frozen duration of Qinghai Lake using satellite passive microwave remote sensing low frequency data, Chin. Sci. Bull., 54 (2009), 2294–2299. https://doi.org/10.1007/s11434-009-0044-3 doi: 10.1007/s11434-009-0044-3
|
| [25] |
L. Feng, J. Liu, T. A. Ali, J. S. Li, J. Li, S. Kuang, Impacts of the decreased freeze-up period on primary production in Qinghai Lake, Int. J. Appl. Earth Obs., 83 (2019), 101915. https://doi.org/10.1016/j.jag.2019.101915 doi: 10.1016/j.jag.2019.101915
|
| [26] |
Z. Jin, C. F. You, Y. Wang, Y. Shi, Hydrological and solute budgets of Lake Qinghai, the largest lake on the Tibetan Plateau, Quatern. Int., 218 (2010), 151–156. https://doi.org/10.1016/j.quaint.2009.11.024 doi: 10.1016/j.quaint.2009.11.024
|
| [27] |
N. Zhang, X. Cao, Q. Xu, X. Huang, U. Herzschuh, Z. Shen, et al., Vegetation change and human-environment interactions in the Qinghai Lake Basin, northeastern Tibetan Plateau, since the last deglaciation, CATENA, 210 (2022), 105892. https://doi.org/10.1016/j.catena.2021.105892 doi: 10.1016/j.catena.2021.105892
|
| [28] |
X. Wang, T. Liang, H Xie, X. Huang, H. Lin, Climate-driven changes in grassland vegetation, snow cover, and lake water of the Qinghai Lake basin, J. Appl. Remote Sens., 10 (2016), 036017. https://doi.org/10.1117/1.JRS.10.036017 doi: 10.1117/1.JRS.10.036017
|
| [29] |
H. Zhang, L. Tian, E. Hasi, D. Zhang, W. Wu, Vegetation-soil dynamics in an alpine desert ecosystem of the Qinghai Lake watershed, northeastern Qinghai-Tibet Plateau, Front. Environ. Sci., 11 (2023), 1119605. https://doi.org/10.3389/fenvs.2023.1119605 doi: 10.3389/fenvs.2023.1119605
|
| [30] |
Y. Cai, J. Zhang, N. Yang, C. Zhang, C. Zhao, H. Long, Human impacts on vegetation exceeded the hydroclimate control 2 ka ago in the Qinghai Lake basin revealed by n-alkanes of loess, Palaeogeogr. Palaeocl., 607 (2022), 111269. https://doi.org/10.1016/j.palaeo.2022.111269 doi: 10.1016/j.palaeo.2022.111269
|
| [31] |
Z. Chen, J. Liu, L. Li, Y. P. Wu, G. Feng, Z. Qian, et al., Effects of climate change on vegetation patterns in Hulun Buir Grassland, Phys. A, 597 (2022), 127275. https://doi.org/10.1016/j.physa.2022.127275 doi: 10.1016/j.physa.2022.127275
|
| [32] |
G. Q. Sun, C. H. Wang, L. L. Chang, Y. P. Wu, L. Li, Z. Jin, Effects of feedback regulation on vegetation patterns in semi-arid environments, Appl. Math. Model., 61 (2018), 200–215. https://doi.org/10.1016/j.apm.2018.04.010 doi: 10.1016/j.apm.2018.04.010
|
| [33] |
J. Liang, C. liu, G. Q. Sun, L. li, L. Zhang, M. Hou, et al., Nonlocal interactions between vegetation induce spatial patterning, Appl. Math. Comput., 428 (2022), 127061. https://doi.org/10.1016/j.amc.2022.127061 doi: 10.1016/j.amc.2022.127061
|
| [34] |
Q. Xue, G. Q. Sun, C. Liu, Z. G. Guo, Z. Jin, Y. P. Wu, et al., Spatiotemporal dynamics of a vegetation model with nonlocal delay in semi-arid environment, Nonlinear Dyn., 99 (2020), 3407–3420. https://doi.org/10.1007/s11071-020-05486-w doi: 10.1007/s11071-020-05486-w
|
| [35] |
M. R. Aguiar, O. E. Sala, M. R. Aguiar, O. E. Sala, Patch structure, dynamics and implications for the functioning of arid ecosystems, Trends Ecol. Evol., 14 (1999), 273–277. https://doi.org/10.1016/S0169-5347(99)01612-2 doi: 10.1016/S0169-5347(99)01612-2
|
| [36] |
C. A. Klausmeier, Regular and irregular patterns in semiarid vegetation, Science, 284 (1999), 1826–1828. https://doi.org/10.1126/science.284.5421.1826 doi: 10.1126/science.284.5421.1826
|
| [37] |
S. K$\acute{e}$fi, M. Rietkerk, G. G. Katul, Vegetation pattern shift as a result of rising atmospheric $CO_2$ in arid ecosystems, Theor. Popul. Biol., 74 (2008), 332–344. https://doi.org/10.1016/j.tpb.2008.09.004 doi: 10.1016/j.tpb.2008.09.004
|
| [38] |
X. Tang, Y. Song, T. Zhang, Turing-Hopf bifurcation analysis of a predator-prey model with herd behavior and cross-diffusion, Nonlinear Dyn., 86 (2016), 73–89. https://doi.org/10.1007/s11071-016-2873-3 doi: 10.1007/s11071-016-2873-3
|
| [39] |
W. Zuo, Y. Song, Stability and bifurcation analysis of a reaction-diffusion equation with distributed delay, Nonlinear Dyn., 79 (2015), 437–454. https://doi.org/10.1007/s11071-014-1677-6 doi: 10.1007/s11071-014-1677-6
|
| [40] |
M. R. Garvie, C. Trenchea, Optimal control of a nutrient-phytoplankton-zooplankton-fish system, SIAM J. Control Optim., 46 (2007), 775–791. https://doi.org/10.1137/050645415 doi: 10.1137/050645415
|
| [41] |
S. Lee, G. Chowell, Exploring optimal control strategies in seasonally varying flu-like epidemics, J. Theor. Biol., 412 (2017), 36–47. https://doi.org/10.1016/j.jtbi.2016.09.023 doi: 10.1016/j.jtbi.2016.09.023
|
| [42] |
L. Chang, S. Gao, Z. Wang, Optimal control of pattern formations for an SIR reaction-diffusion epidemic model, J. Theor. Biol., 536 (2022), 111003. https://doi.org/10.1016/j.jtbi.2022.111003 doi: 10.1016/j.jtbi.2022.111003
|
| [43] |
L. Chang, W. Gong, Z. Jin, G. Q. Sun, Sparse optimal control of pattern formations for an SIR reaction-diffusion epidemic model, SIAM J. Appl. Math., 82 (2022), 1764–1790. https://doi.org/10.1137/22M1472127 doi: 10.1137/22M1472127
|
| [44] |
W. Choi, E. Shim, Optimal strategies for social distancing and testing to control COVID-19, J. Theor. Biol., 512 (2021), 110568. https://doi.org/10.1016/j.jtbi.2020.110568 doi: 10.1016/j.jtbi.2020.110568
|
| [45] |
S. Kim, J. Lee, E. Jung, Mathematical model of transmission dynamics and optimal control strategies for 2009 A/H1N1 influenza in the Republic of Korea, J. Theor. Biol., 412 (2017), 74–85. https://doi.org/10.1016/j.jtbi.2016.09.025 doi: 10.1016/j.jtbi.2016.09.025
|
| [46] |
K. E. Taylor, R. J. Stouffer, G. A. Meehl, An overview of CMIP5 and the experiment design, Bull. Amer. Meteorol. Soc., 93 (2012), 485–498. https://doi.org/10.1175/BAMS-D-11-00094.1 doi: 10.1175/BAMS-D-11-00094.1
|
| [47] |
K. Calvin, B. Bond-Lamberty, L. Clarke, J. Edmonds, J. Eom, C. Hartin, et al., The SSP4: A world of deepening inequality, Global Environ. Chang, 42 (2017), 284–296. https://doi.org/10.1016/j.gloenvcha.2016.06.010 doi: 10.1016/j.gloenvcha.2016.06.010
|
| [48] |
T. Zhao, L. Chen, Z. Ma, Simulation of historical and projected climate change in arid and semiarid areas by CMIP5 models, Chin. Sci. Bull., 59 (2014), 412–429. https://doi.org/10.1007/s11434-013-0003-x doi: 10.1007/s11434-013-0003-x
|
| [49] |
J. von Hardenberg, E. Meron, M. Shachak, Y. Zarmi, Diversity of vegetation patterns and desertification, Phys. Rev. Lett., 87 (2001), 198101. https://doi.org/10.1103/PhysRevLett.87.198101 doi: 10.1103/PhysRevLett.87.198101
|






Figures(7) / Tables(2)

Juan Liang, Huilian Ma, Huanqing Yang, Zunguang Guo. Response of vegetation pattern to climate change based on dynamical model: Case of Qinghai Lake, China[J]. AIMS Mathematics, 2024, 9(1): 2500-2517. doi: 10.3934/math.2024123







 DownLoad:
DownLoad: