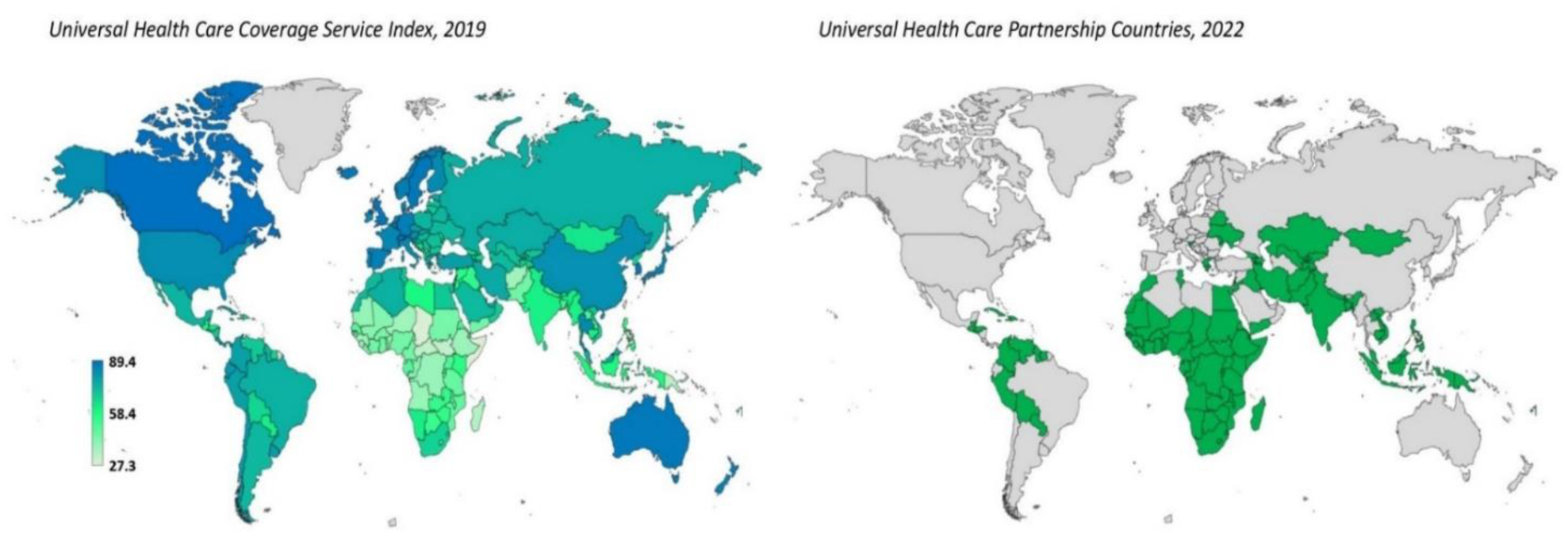
In November 2022, the global population had officially crossed eight billion. It has long been recognized that socioeconomic or health-related problems in the community always accompany an uncontrolled population expansion. International calls have been made regarding lack of universal health coverage, an insufficient supply of healthcare providers, the burden of noncommunicable disease, population aging and the difficulty in obtaining safe drinking water and food. The present health policy paper discusses how to conquer these crowded world issues, including (1) promoting government and international organization participation in providing appropriate infrastructure, funding and distribution to assist people's health and well-being; (2) shifting health program towards a more preventive approach and (3) reducing inequalities, particularly for the marginalized, isolated and underrepresented population. These fundamental principles of health policy delivery as a response to an increasingly crowded world and its challenges are crucial for reducing the burden associated with excessive healthcare costs, decreased productivity and deteriorating environmental quality.
Citation: Nityanand Jain, Islam Kourampi, Tungki Pratama Umar, Zahra Rose Almansoor, Ayush Anand, Mohammad Ebad Ur Rehman, Shivani Jain, Aigars Reinis. Global population surpasses eight billion: Are we ready for the next billion?[J]. AIMS Public Health, 2023, 10(4): 849-866. doi: 10.3934/publichealth.2023056
In November 2022, the global population had officially crossed eight billion. It has long been recognized that socioeconomic or health-related problems in the community always accompany an uncontrolled population expansion. International calls have been made regarding lack of universal health coverage, an insufficient supply of healthcare providers, the burden of noncommunicable disease, population aging and the difficulty in obtaining safe drinking water and food. The present health policy paper discusses how to conquer these crowded world issues, including (1) promoting government and international organization participation in providing appropriate infrastructure, funding and distribution to assist people's health and well-being; (2) shifting health program towards a more preventive approach and (3) reducing inequalities, particularly for the marginalized, isolated and underrepresented population. These fundamental principles of health policy delivery as a response to an increasingly crowded world and its challenges are crucial for reducing the burden associated with excessive healthcare costs, decreased productivity and deteriorating environmental quality.

| [1] | The Lancet.A world of 7 billion people. Lancet (2011) 378: 1527. https://doi.org/10.1016/S0140-6736(11)61658-9 |
| [2] | Department of Economic and Social AffairsWorld population to reach 8 billion on 15 November 2022 (2022). Available from: https://www.un.org/en/desa/world-population-reach-8-billion-15-november-2022. |
| [3] | World Health OrganizationUniversal health coverage (2021). Available from: https://www.who.int/news-room/fact-sheets/detail/universal-health-coverage-(uhc). |
| [4] |
Odoch WD, Senkubuge F, Masese AB, et al. (2023) A critical review of literature on health financing reforms in Uganda-progress, challenges and opportunities for achieving UHC. Afr Health Sci 23: 736-746. https://doi.org/10.4314/ahs.v23i1.78 
|
| [5] | World Health OrganizationGlobal oral health status report: Towards universal health coverage for oral health by 2030 (2022). Available from: https://www.paho.org/en/documents/global-oral-health-status-report-towards-universal-health-coverage-oral-health-2030 |
| [6] | (2022) Assistant Secretary for Public Affairs (ASPA)Ahead of open enrollment, new HHS Report shows major gains in marketplace enrollment among Black and Latino populations last year. USA: HHS Press Office. Available from: https://www.hhs.gov/about/news/2022/10/25/ahead-of-open-enrollment-new-hhs-report-shows-major-gains-in-marketplace-enrollment-among-black-and-latino-populations-last-year.html |
| [7] |
Tan X, Liu X, Shao H (2017) Healthy China 2030: A vision for health care. Value Health Reg Issues 12: 112-114. https://doi.org/10.1016/j.vhri.2017.04.001
|
| [8] |
Angell BJ, Prinja S, Gupt A, et al. (2019) The Ayushman Bharat Pradhan Mantri Jan Arogya Yojana and the path to universal health coverage in India: Overcoming the challenges of stewardship and governance. PLoS Med 16: e1002759. https://doi.org/10.1371/journal.pmed.1002759
|
| [9] | General Assembly.Resolution adopted by the general assembly on 10 October 2019-A/RES/74/2 “Political declaration of the high-level meeting on universal health coverage”. 74th United Nations GA Session (2022) . Available from: https://documents-dds-ny.un.org/doc/UNDOC/GEN/N19/311/84/PDF/N1931184.pdf?OpenElement |
| [10] |
Kumar R, Pal R (2018) India achieves WHO recommended doctor population ratio: A call for paradigm shift in public health discourse!. J Family Med Prim Care 7: 841-844. https://doi.org/10.4103/jfmpc.jfmpc_218_18
|
| [11] | Schwartz E (2022) The global health care worker shortage: 10 Numbers to note. Project Hope . Available from: https://www.projecthope.org/the-global-health-worker-shortage-10-numbers-to-note/04/2022/ |
| [12] |
Boniol M, Kunjumen T, Nair TS, et al. (2022) The global health workforce stock and distribution in 2020 and 2030: a threat to equity and ‘universal’ health coverage?. BMJ Glob Health 7: e009316. https://doi.org/10.1136/bmjgh-2022-009316
|
| [13] |
Owens B (2018) If Canada needs more doctors, why hasn't medical school enrolment increased?. CMAJ 190: E1266-E1267. https://doi.org/10.1503/cmaj.109-5649
|
| [14] |
McGrail MR, O'Sullivan BG (2021) Increasing doctors working in specific rural regions through selection from and training in the same region: National evidence from Australia. Hum Resour Health 19: 132. https://doi.org/10.1186/s12960-021-00678-w
|
| [15] |
Adovor E, Czaika M, Docquier F, et al. (2021) Medical brain drain: How many, where and why?. J Health Econ 76: 102409. https://doi.org/10.1016/j.jhealeco.2020.102409
|
| [16] |
Newman C (2014) Time to address gender discrimination and inequality in the health workforce. Hum Resour Health 12: 25. https://doi.org/10.1186/1478-4491-12-25
|
| [17] |
Hay K, McDougal L, Percival V, et al. (2019) Disrupting gender norms in health systems: making the case for change. Lancet 393: 2535-2549. https://doi.org/10.1016/S0140-6736(19)30648-8
|
| [18] |
Wu LT, Low MM, Tan KK, et al. (2015) Why not nursing? A systematic review of factors influencing career choice among healthcare students. Int Nurs Rev 62: 547-562. https://doi.org/10.1111/inr.12220
|
| [19] |
Hasebrook J, Hahnenkamp K, Buhre WFFA, et al. (2017) Medicine goes female: Protocol for improving career options of females and working conditions for researching physicians in clinical medical research by organizational transformation and participatory design. JMIR Res Protoc 6: e152. https://doi.org/10.2196/resprot.7632
|
| [20] |
Murthy VH (2022) Confronting health worker burnout and well-being. N Engl J Med 387: 577-579. https://doi.org/10.1056/NEJMp2207252
|
| [21] |
Sinsky CA, Brown RL, Stillman MJ, et al. (2021) COVID-related stress and work intentions in a sample of US health care workers. Mayo Clin Proc Innov Qual Outcomes 5: 1165-1173. https://doi.org/10.1016/j.mayocpiqo.2021.08.007
|
| [22] | World Health OrganizationNoncommunicable diseases (2022). Available from: https://www.who.int/news-room/fact-sheets/detail/noncommunicable-diseases |
| [23] | World Health OrganizationPreventing chronic diseases: A vital investment: WHO global report (2005). Available from: https://apps.who.int/iris/handle/10665/43314 |
| [24] |
Mirelman AJ, Rose S, Khan JA, et al. (2016) The relationship between non-communicable disease occurrence and poverty-evidence from demographic surveillance in Matlab, Bangladesh. Health Policy Plan 31: 785-792. https://doi.org/10.1093/heapol/czv134
|
| [25] | Shiferaw F, Letebo M, Feleke Y, et al. (2019) Non-communicable diseases in Ethiopia: Policy and strategy gaps in the reduction of behavioral risk factors. Ethiop J Health Development 33: 260-267. |
| [26] | Albelbeisi AH, Albelbeisi A, El Bilbeisi AH, et al. (2021) Public sector capacity to prevent and control of noncommunicable diseases in twelve low- and middle-income countries based on WHO-PEN standards: A systematic review. Health Serv Insights 14: 1178632920986233. https://doi.org/10.1177/1178632920986233 |
| [27] |
Tripathy JP (2018) Research priorities in non-communicable diseases in developing countries: Time to go beyond prevalence studies. Public Health Action 8: 98-99. https://doi.org/10.5588/pha.18.0023
|
| [28] |
Mendis S, Johnston SC, Fan W, et al. (2010) Cardiovascular risk management and its impact on hypertension control in primary care in low-resource settings: A cluster-randomized trial. Bull World Health Organ 88: 412-419. https://doi.org/10.2471/BLT.08.062364
|
| [29] | Zhao S, Zhang Y, Xiao AY, et al. (2023) Key factors associated with quality of postnatal care: A pooled analysis of 23 countries. E Clinical Medicine 62: 102090. https://doi.org/10.1016/j.eclinm.2023.102090 |
| [30] |
Fritz M, Fromell H (2022) How to dampen the surge of non-communicable diseases in Southeast Asia: insights from a systematic review and meta-analysis. Health Policy Plan 37: 152-167. https://doi.org/10.1093/heapol/czab138
|
| [31] |
Santos JA, Tekle D, Rosewarne E, et al. (2021) A systematic review of salt reduction initiatives around the world: A midterm evaluation of progress towards the 2025 global non-communicable diseases salt reduction target. Adv Nutr 12: 1768-1780. https://doi.org/10.1093/advances/nmab008
|
| [32] | World Health OrganizationWHO launches Signature Initiative to reduce cardiovascular disease through salt reduction (2022). Available from: https://www.who.int/europe/news-room/events/item/2022/12/09/default-calendar/who-launches-signature-initiative-to-reduce-cardiovascular-disease-through-salt-reduction |
| [33] |
Mohammed S, Afaya A, Abukari AS (2023) Reading, singing, and storytelling: The impact of caregiver-child interaction and child access to books and preschool on early childhood development in Ghana. Sci Rep 13: 13751. https://doi.org/10.1038/s41598-023-38439-5
|
| [34] |
Eriksson MK, Hagberg L, Lindholm L, et al. (2010) Quality of life and cost-effectiveness of a 3-year trial of lifestyle intervention in primary health care. Arch Intern Med 170: 1470-1479. https://doi.org/10.1001/archinternmed.2010.301
|
| [35] |
Romé A, Persson U, Ekdahl C, et al. (2009) Physical activity on prescription (PAP): costs and consequences of a randomized, controlled trial in primary healthcare. Scand J Prim Health Care 27: 216-222. https://doi.org/10.3109/02813430903438734
|
| [36] | World Health OrganizationAgeing and health (2022). Available from: https://www.who.int/news-room/fact-sheets/detail/ageing-and-health |
| [37] |
Cheng X, Yang Y, Schwebel DC, et al. (2020) Population ageing and mortality during 1990–2017: A global decomposition analysis. PLoS Med 17: e1003138. https://doi.org/10.1371/journal.pmed.1003138
|
| [38] |
Tipirneni R, Solway E, Malani P, et al. (2020) Health Insurance Affordability Concerns and Health Care Avoidance Among US Adults Approaching Retirement. JAMA Netw Open 3: e1920647. https://doi.org/10.1001/jamanetworkopen.2019.20647
|
| [39] |
Gabrani J, Schindler C, Wyss K (2022) Out of pocket payments and access to NCD medication in two regions in Albania. PLoS One 17: e0272221. https://doi.org/10.1371/journal.pone.0272221
|
| [40] |
Hock H, Weil DN (2012) On the dynamics of the age structure, dependency, and consumption. J Popul Econ 25: 10.1007/s00148-011-0372-x. https://doi.org/10.1007/s00148-011-0372-x
|
| [41] |
Malik C, Khanna S, Jain Y, et al. (2021) Geriatric population in India: Demography, vulnerabilities, and healthcare challenges. J Family Med Prim Care 10: 72-76. https://doi.org/10.4103/jfmpc.jfmpc_1794_20
|
| [42] |
Murdaca G, Orsi A, Spanò F, et al. (2014) Influenza and pneumococcal vaccinations of patients with systemic lupus erythematosus: current views upon safety and immunogenicity. Autoimmun Rev 13: 75-84. https://doi.org/10.1016/j.autrev.2013.07.007
|
| [43] |
Murdaca G, Orsi A, Spanò F, et al. (2016) Vaccine-preventable infections in Systemic Lupus Erythematosus. Hum Vaccin Immunother 12: 632-643. Https://doi.org/10.1080/21645515.2015.1107685
|
| [44] |
Fulmer T, Reuben DB, Auerbach J, et al. (2021) Actualizing better health and health care for older adults. Health Aff (Millwood) 40: 219-225. https://doi.org/10.1377/hlthaff.2020.01470
|
| [45] | Indriani D (2020) Efektivitas Implementasi Badan Penyelenggaraan Jaminan Sosial (BPJS) Serta Dampaknya Terhadap Kesehatan Dan Ekonomi Masyarakat Tahun 2009–2013 Dan 2015–2019. J Ris Mhs 1: 76-85. https://doi.org/10.23969/brainy.v1i1.11 |
| [46] |
Combi C, Pozzani G, Pozzi G (2016) Telemedicine for developing countries. A survey and some design issues. Appl Clin Inform 7: 1025-1050. https://doi.org/10.4338/ACI-2016-06-R-0089
|
| [47] | World Health OrganizationTelemedicine has clear benefits for patients in European countries, new study shows. Europe World Health Organization (2022). Available from: https://www.who.int/europe/news/item/31-10-2022-telemedicine-has-clear-benefits-for-patients-in-european-countries--new-study-shows |
| [48] |
Fernandez-Lazaro CI, García-González JM, Adams DP, et al. (2019) Adherence to treatment and related factors among patients with chronic conditions in primary care: A cross-sectional study. BMC Fam Pract 20: 132. https://doi.org/10.1186/s12875-019-1019-3
|
| [49] |
Tkatch R, Musich S, MacLeod S, et al. (2016) Population health management for older adults: Review of interventions for promoting successful aging across the health continuum. Gerontol Geriatr Med 2: 2333721416667877. https://doi.org/10.1177/2333721416667877
|
| [50] |
Macinko J, Cristina Drumond Andrade F, Bof de Andrade F, et al. (2020) Universal health coverage: Are older adults being left behind? Evidence from aging cohorts in twenty-three countries. Health Aff (Millwood) 39: 1951-1960. https://doi.org/10.1377/hlthaff.2019.01570
|
| [51] |
Okamoto S, Komamura K (2022) Towards universal health coverage in the context of population ageing: a narrative review on the implications from the long-term care system in Japan. Arch Public Health 80: 210. https://doi.org/10.1186/s13690-022-00970-8
|
| [52] |
Jacques E, Suuronen EJ (2020) The progression of regenerative medicine and its impact on therapy translation. Clin Transl Sci 13: 440-450. https://doi.org/10.1111/cts.12736
|
| [53] |
Mao AS, Mooney DJ (2015) Regenerative medicine: Current therapies and future directions. Proc Natl Acad Sci USA 112: 14452-14459. https://doi.org/10.1073/pnas.1508520112
|
| [54] |
Lovell-Badge R, Anthony E, Barker RA, et al. (2021) ISSCR Guidelines for stem cell research and clinical translation: The 2021 update. Stem Cell Reports 16: 1398-1408. https://doi.org/10.1016/j.stemcr.2021.05.012
|
| [55] |
Russa AD (2019) Stem cells: Prospects and potential applications in Tanzania: A review. Tanzan J Health Res 21: 1-8. https://doi.org/10.4314/thrb.v21i2.8
|
| [56] |
Ricci F (2012) Social implications of malaria and their relationships with poverty. Mediterr J Hematol Infect Dis 4: e2012048. https://doi.org/10.4084/MJHID.2012.048
|
| [57] |
Nguyen-Tien T, Do DC, Le XL, et al. (2021) Risk factors of dengue fever in an urban area in Vietnam: A case-control study. BMC Public Health 21: 664. https://doi.org/10.1186/s12889-021-10687-y
|
| [58] | Neiderud CJ (2015) How urbanization affects the epidemiology of emerging infectious diseases. Infect Ecol Epidemiol 5: 27060. https://doi.org/10.3402/iee.v5.27060 |
| [59] | Carballo M, Maclean EC, Gudumac I, et al. (2016) Hepatitis C and migration: A public health challenge. J Fam Med 3: 1065. |
| [60] | World Economic Forum (WEF)The art and science of eliminating Hepatitis: Egypt's experience (2022). Available from: https://www.weforum.org/whitepapers/the-art-and-science-of-eliminating-hepatitis-egypt-s-experience/ |
| [61] |
Boretti A, Rosa L (2019) Reassessing the projections of the World Water Development Report. NPJ Clean Water 2: 15. https://doi.org/10.1038/s41545-019-0039-9
|
| [62] |
Tarrass F, Benjelloun M (2012) The effects of water shortages on health and human development. Perspect Public Health 132: 240-244. https://doi.org/10.1177/1757913910391040
|
| [63] |
Ahmed T, Zounemat-Kermani M, Scholz M (2020) Climate change, water quality and water-related challenges: A review with focus on Pakistan. Int J Environ Res Public Health 17: 8518. https://doi.org/10.3390/ijerph17228518
|
| [64] |
Strunz EC, Addiss DG, Stocks ME, et al. (2014) Water, sanitation, hygiene, and soil-transmitted helminth infection: A systematic review and meta-analysis. PLoS Med 11: e1001620. https://doi.org/10.1371/journal.pmed.1001620
|
| [65] | Garn JV, Wilkers JL, Meehan AA, et al. (2022) Interventions to improve water, sanitation, and hygiene for preventing soil-transmitted helminth infection. Cochrane Database Syst Rev 6: CD012199. https://doi.org/10.1002/14651858.CD012199.pub2 |
| [66] |
van Dijk M, Morley T, Rau ML, et al. (2021) A meta-analysis of projected global food demand and population at risk of hunger for the period 2010–2050. Nat Food 2: 494-501. https://doi.org/10.1038/s43016-021-00322-9
|
| [67] | Cartín-Rojas A (2012) Transboundary animal diseases and international trade. International Trade from Economic and Policy Perspective [online] . London: IntechOpen. https://doi.org/10.5772/48151 |
| [68] | WHO Global Centre for Traditional MedicineCatalysing ancient wisdom and modern science for the health of people and the planet (2022). Available from: https://www.who.int/initiatives/who-global-centre-for-traditional-medicine |
| [69] | Madamombe I (2006) Traditional healers boost primary health care. Africa Renewal (United Nations) . Available from: https://www.un.org/africarenewal/magazine/january-2006/traditional-healers-boost-primary-health-care |
| [70] | Health in HarmonyImpact Report 2021 (2022). Available from: https://healthinharmony.org/wp-content/uploads/2021-HIH-Impact-Report_Final_small.pdf |



Figures(4)

Nityanand Jain, Islam Kourampi, Tungki Pratama Umar, Zahra Rose Almansoor, Ayush Anand, Mohammad Ebad Ur Rehman, Shivani Jain, Aigars Reinis. Global population surpasses eight billion: Are we ready for the next billion?[J]. AIMS Public Health, 2023, 10(4): 849-866. doi: 10.3934/publichealth.2023056







 DownLoad:
DownLoad: