Citation: Xin Nian, Yasuhiro Nagai, Cameron Jeffers, Kara N. Maxwell, Hongtao Zhang. Dietary influence on estrogens and cytokines in breast cancer[J]. AIMS Molecular Science, 2017, 4(3): 252-270. doi: 10.3934/molsci.2017.3.252

| [1] | Howlader N, Noone A, Krapcho M, et al. (2015) SEER Cancer Statistics Review, 1975–2012, National Cancer Institute. Bethesda, MD. 2015. |
| [2] |
Tomasetti C, Li L, Vogelstein B (2017) Stem cell divisions, somatic mutations, cancer etiology, and cancer prevention. Science 355: 1330-1334. doi: 10.1126/science.aaf9011

|
| [3] | Cancer Research UK, Statistics on preventable cancers. 2017. Available from: http://www.cancerresearchuk.org/health-professional/cancer-statistics/risk/preventable-cancers#heading-One |
| [4] |
Colditz GA, Hankinson SE (2005) The Nurses' Health Study: lifestyle and health among women. Nat Rev Cancer 5: 388-396. doi: 10.1038/nrc1608
|
| [5] |
Boeke CE, Eliassen AH, Chen WY, et al. (2014) Dietary fat intake in relation to lethal breast cancer in two large prospective cohort studies. Breast Cancer Res Treat 146: 383-392. doi: 10.1007/s10549-014-3005-8
|
| [6] |
Farvid MS, Cho E, Chen WY, et al. (2014) Premenopausal dietary fat in relation to pre- and post-menopausal breast cancer. Breast Cancer Res Treat 145: 255-265. doi: 10.1007/s10549-014-2895-9
|
| [7] |
Sasaki S, Horacsek M, Kesteloot H (1993) An ecological study of the relationship between dietary fat intake and breast cancer mortality. Prev Med 22: 187-202. doi: 10.1006/pmed.1993.1016
|
| [8] |
Chlebowski RT, Blackburn GL, Thomson CA, et al. (2006) Dietary fat reduction and breast cancer outcome: interim efficacy results from the Women's Intervention Nutrition Study. J Natl Cancer Inst 98: 1767-1776. doi: 10.1093/jnci/djj494
|
| [9] | Chlebowski RT (2013) Nutrition and physical activity influence on breast cancer incidence and outcome. Breast 22 Suppl 2: S30-37. |
| [10] |
Pierce JP, Natarajan L, Caan BJ, et al. (2007) Influence of a diet very high in vegetables, fruit, and fiber and low in fat on prognosis following treatment for breast cancer: the Women's Healthy Eating and Living (WHEL) randomized trial. JAMA 298: 289-298. doi: 10.1001/jama.298.3.289
|
| [11] | Sieri S, Chiodini P, Agnoli C, et al. (2014) Dietary fat intake and development of specific breast cancer subtypes. J Natl Cancer Inst 106. |
| [12] |
Cao Y, Hou L, Wang W (2016) Dietary total fat and fatty acids intake, serum fatty acids and risk of breast cancer: A meta-analysis of prospective cohort studies. Int J Cancer 138: 1894-1904. doi: 10.1002/ijc.29938
|
| [13] |
Bagga D, Anders KH, Wang HJ, et al. (2002) Long-chain n-3-to-n-6 polyunsaturated fatty acid ratios in breast adipose tissue from women with and without breast cancer. Nutr Cancer 42: 180-185. doi: 10.1207/S15327914NC422_5
|
| [14] |
Murff HJ, Shu XO, Li H, et al. (2011) Dietary polyunsaturated fatty acids and breast cancer risk in Chinese women: a prospective cohort study. Int J Cancer 128: 1434-1441. doi: 10.1002/ijc.25703
|
| [15] |
Gago-Dominguez M, Yuan JM, Sun CL, et al. (2003) Opposing effects of dietary n-3 and n-6 fatty acids on mammary carcinogenesis: The Singapore Chinese Health Study. Br J Cancer 89: 1686-1692. doi: 10.1038/sj.bjc.6601340
|
| [16] |
Khankari NK, Bradshaw PT, Steck SE, et al. (2015) Dietary intake of fish, polyunsaturated fatty acids, and survival after breast cancer: A population-based follow-up study on Long Island, New York. Cancer 121: 2244-2252. doi: 10.1002/cncr.29329
|
| [17] |
Wakai K, Tamakoshi K, Date C, et al. (2005) Dietary intakes of fat and fatty acids and risk of breast cancer: a prospective study in Japan. Cancer Sci 96: 590-599. doi: 10.1111/j.1349-7006.2005.00084.x
|
| [18] |
Kiyabu GY, Inoue M, Saito E, et al. (2015) Fish, n - 3 polyunsaturated fatty acids and n - 6 polyunsaturated fatty acids intake and breast cancer risk: The Japan Public Health Center-based prospective study. Int J Cancer 137: 2915-2926. doi: 10.1002/ijc.29672
|
| [19] |
Bagga D, Wang L, Farias-Eisner R, et al. (2003) Differential effects of prostaglandin derived from omega-6 and omega-3 polyunsaturated fatty acids on COX-2 expression and IL-6 secretion. Proc Natl Acad Sci U S A 100: 1751-1756. doi: 10.1073/pnas.0334211100
|
| [20] | Chenais B, Blanckaert V (2012) The janus face of lipids in human breast cancer: how polyunsaturated Fatty acids affect tumor cell hallmarks. Int J Breast Cancer 2012: 712536. |
| [21] |
Schwingshackl L, Hoffmann G (2016) Does a Mediterranean-Type Diet Reduce Cancer Risk? Curr Nutr Rep 5: 9-17. doi: 10.1007/s13668-015-0141-7
|
| [22] |
van den Brandt PA, Schulpen M (2017) Mediterranean diet adherence and risk of postmenopausal breast cancer: results of a cohort study and meta-analysis. Int J Cancer 140: 2220-2231. doi: 10.1002/ijc.30654
|
| [23] |
Pot GK, Stephen AM, Dahm CC, et al. (2014) Dietary patterns derived with multiple methods from food diaries and breast cancer risk in the UK Dietary Cohort Consortium. Eur J Clin Nutr 68: 1353-1358. doi: 10.1038/ejcn.2014.135
|
| [24] |
Toledo E, Salas-Salvado J, Donat-Vargas C, et al. (2015) Mediterranean Diet and Invasive Breast Cancer Risk Among Women at High Cardiovascular Risk in the PREDIMED Trial: A Randomized Clinical Trial. JAMA Intern Med 175: 1752-1760. doi: 10.1001/jamainternmed.2015.4838
|
| [25] | Farvid MS, Chen WY, Michels KB, et al. (2016) Fruit and vegetable consumption in adolescence and early adulthood and risk of breast cancer: population based cohort study. BMJ 353: i2343. |
| [26] |
Liu Y, Colditz GA, Cotterchio M, et al. (2014) Adolescent dietary fiber, vegetable fat, vegetable protein, and nut intakes and breast cancer risk. Breast Cancer Res Treat 145: 461-470. doi: 10.1007/s10549-014-2953-3
|
| [27] | Dam MK, Hvidtfeldt UA, Tjonneland A, et al. (2016) Five year change in alcohol intake and risk of breast cancer and coronary heart disease among postmenopausal women: prospective cohort study. BMJ 353: i2314. |
| [28] |
Dong JY, Qin LQ (2011) Dietary glycemic index, glycemic load, and risk of breast cancer: meta-analysis of prospective cohort studies. Breast Cancer Res Treat 126: 287-294. doi: 10.1007/s10549-011-1343-3
|
| [29] |
Inoue-Choi M, Sinha R, Gierach GL, et al. (2016) Red and processed meat, nitrite, and heme iron intakes and postmenopausal breast cancer risk in the NIH-AARP Diet and Health Study. Int J Cancer 138: 1609-1618. doi: 10.1002/ijc.29901
|
| [30] |
Touvier M, Fassier P, His M, et al. (2015) Cholesterol and breast cancer risk: a systematic review and meta-analysis of prospective studies. Br J Nutr 114: 347-357. doi: 10.1017/S000711451500183X
|
| [31] |
Tornberg SA, Holm LE, Carstensen JM (1988) Breast cancer risk in relation to serum cholesterol, serum beta-lipoprotein, height, weight, and blood pressure. Acta Oncol 27: 31-37. doi: 10.3109/02841868809090315
|
| [32] | Vatten LJ, Foss OP (1990) Total serum cholesterol and triglycerides and risk of breast cancer: a prospective study of 24,329 Norwegian women. Cancer Res 50: 2341-2346. |
| [33] | Furberg AS, Jasienska G, Bjurstam N, et al. (2005) Metabolic and hormonal profiles: HDL cholesterol as a plausible biomarker of breast cancer risk. The Norwegian EBBA Study. Cancer Epidemiol Biomarkers Prev 14: 33-40. |
| [34] |
Ha M, Sung J, Song YM (2009) Serum total cholesterol and the risk of breast cancer in postmenopausal Korean women. Cancer Causes Control 20: 1055-1060. doi: 10.1007/s10552-009-9301-7
|
| [35] |
Kitahara CM, Berrington de Gonzalez A, Freedman ND, et al. (2011) Total cholesterol and cancer risk in a large prospective study in Korea. J Clin Oncol 29: 1592-1598. doi: 10.1200/JCO.2010.31.5200
|
| [36] |
Li C, Yang L, Zhang D, et al. (2016) Systematic review and meta-analysis suggest that dietary cholesterol intake increases risk of breast cancer. Nutr Res 36: 627-635. doi: 10.1016/j.nutres.2016.04.009
|
| [37] |
McDonnell DP, Park S, Goulet MT, et al. (2014) Obesity, cholesterol metabolism, and breast cancer pathogenesis. Cancer Res 74: 4976-4982. doi: 10.1158/0008-5472.CAN-14-1756
|
| [38] |
Reeves GK, Pirie K, Beral V, et al. (2007) Cancer incidence and mortality in relation to body mass index in the Million Women Study: cohort study. BMJ 335: 1134. doi: 10.1136/bmj.39367.495995.AE
|
| [39] |
Renehan AG, Zwahlen M, Egger M (2015) Adiposity and cancer risk: new mechanistic insights from epidemiology. Nat Rev Cancer 15: 484-498. doi: 10.1038/nrc3967
|
| [40] |
Baumgartner KB, Hunt WC, Baumgartner RN, et al. (2004) Association of body composition and weight history with breast cancer prognostic markers: divergent pattern for Hispanic and non-Hispanic White women. Am J Epidemiol 160: 1087-1097. doi: 10.1093/aje/kwh313
|
| [41] |
Harris HR, Willett WC, Terry KL, et al. (2011) Body fat distribution and risk of premenopausal breast cancer in the Nurses' Health Study II. J Natl Cancer Inst 103: 273-278. doi: 10.1093/jnci/djq500
|
| [42] |
Jiralerspong S, Goodwin PJ (2016) Obesity and Breast Cancer Prognosis: Evidence, Challenges, and Opportunities. J Clin Oncol 34: 4203-4216. doi: 10.1200/JCO.2016.68.4480
|
| [43] |
Arce-Salinas C, Aguilar-Ponce JL, Villarreal-Garza C, et al. (2014) Overweight and obesity as poor prognostic factors in locally advanced breast cancer patients. Breast Cancer Res Treat 146: 183-188. doi: 10.1007/s10549-014-2977-8
|
| [44] |
Printz C (2014) Obesity associated with higher mortality in women with ER-positive breast cancer. Cancer 120: 3267. doi: 10.1002/cncr.29079
|
| [45] |
Ottobelli Chielle E, de Souza WM, da Silva TP, et al. (2016) Adipocytokines, inflammatory and oxidative stress markers of clinical relevance altered in young overweight/obese subjects. Clin Biochem 49: 548-553. doi: 10.1016/j.clinbiochem.2016.01.003
|
| [46] |
Bulun SE, Chen D, Moy I, et al. (2012) Aromatase, breast cancer and obesity: a complex interaction. Trends Endocrinol Metab 23: 83-89. doi: 10.1016/j.tem.2011.10.003
|
| [47] |
Creighton CJ, Sada YH, Zhang Y, et al. (2012) A gene transcription signature of obesity in breast cancer. Breast Cancer Res Treat 132: 993-1000. doi: 10.1007/s10549-011-1595-y
|
| [48] |
Perks CM, Holly JM (2011) Hormonal mechanisms underlying the relationship between obesity and breast cancer. Endocrinol Metab Clin North Am 40: 485-507, vii. doi: 10.1016/j.ecl.2011.05.010
|
| [49] |
Vadgama JV, Wu Y, Datta G, et al. (1999) Plasma insulin-like growth factor-I and serum IGF-binding protein 3 can be associated with the progression of breast cancer, and predict the risk of recurrence and the probability of survival in African-American and Hispanic women. Oncology 57: 330-340. doi: 10.1159/000012052
|
| [50] |
Kaaks R, Lundin E, Rinaldi S, et al. (2002) Prospective study of IGF-I, IGF-binding proteins, and breast cancer risk, in northern and southern Sweden. Cancer Causes Control 13: 307-316. doi: 10.1023/A:1015270324325
|
| [51] |
Sugumar A, Liu YC, Xia Q, et al. (2004) Insulin-like growth factor (IGF)-I and IGF-binding protein 3 and the risk of premenopausal breast cancer: a meta-analysis of literature. Int J Cancer 111: 293-297. doi: 10.1002/ijc.20253
|
| [52] |
Renehan AG, Harvie M, Howell A (2006) Insulin-like growth factor (IGF)-I, IGF binding protein-3, and breast cancer risk: eight years on. Endocr Relat Cancer 13: 273-278. doi: 10.1677/erc.1.01219
|
| [53] |
Endogenous H, Breast Cancer Collaborative G, Key TJ, et al. (2010) Insulin-like growth factor 1 (IGF1), IGF binding protein 3 (IGFBP3), and breast cancer risk: pooled individual data analysis of 17 prospective studies. Lancet Oncol 11: 530-542. doi: 10.1016/S1470-2045(10)70095-4
|
| [54] |
Rinaldi S, Peeters PH, Berrino F, et al. (2006) IGF-I, IGFBP-3 and breast cancer risk in women: The European Prospective Investigation into Cancer and Nutrition (EPIC). Endocr Relat Cancer 13: 593-605. doi: 10.1677/erc.1.01150
|
| [55] |
Allen NE, Roddam AW, Allen DS, et al. (2005) A prospective study of serum insulin-like growth factor-I (IGF-I), IGF-II, IGF-binding protein-3 and breast cancer risk. Br J Cancer 92: 1283-1287. doi: 10.1038/sj.bjc.6602471
|
| [56] | Rinaldi S, Kaaks R, Zeleniuch-Jacquotte A, et al. (2005) Insulin-like growth factor-I, IGF binding protein-3, and breast cancer in young women: a comparison of risk estimates using different peptide assays. Cancer Epidemiol Biomarkers Prev 14: 48-52. |
| [57] |
Schairer C, Hill D, Sturgeon SR, et al. (2004) Serum concentrations of IGF-I, IGFBP-3 and c-peptide and risk of hyperplasia and cancer of the breast in postmenopausal women. Int J Cancer 108: 773-779. doi: 10.1002/ijc.11624
|
| [58] |
Signori C, DuBrock C, Richie JP, et al. (2012) Administration of omega-3 fatty acids and Raloxifene to women at high risk of breast cancer: interim feasibility and biomarkers analysis from a clinical trial. Eur J Clin Nutr 66: 878-884. doi: 10.1038/ejcn.2012.60
|
| [59] |
Kabat GC, Kim M, Caan BJ, et al. (2009) Repeated measures of serum glucose and insulin in relation to postmenopausal breast cancer. Int J Cancer 125: 2704-2710. doi: 10.1002/ijc.24609
|
| [60] | Goodwin PJ, Ennis M, Pritchard KI, et al. (2012) Insulin- and obesity-related variables in early-stage breast cancer: correlations and time course of prognostic associations. J Clin Oncol 30: 164-171. |
| [61] | Sun W, Lu J, Wu S, et al. (2016) Association of insulin resistance with breast, ovarian, endometrial and cervical cancers in non-diabetic women. Am J Cancer Res 6: 2334-2344. |
| [62] |
Lee JO, Kim N, Lee HJ, et al. (2016) Resistin, a fat-derived secretory factor, promotes metastasis of MDA-MB-231 human breast cancer cells through ERM activation. Sci Rep 6: 18923. doi: 10.1038/srep18923
|
| [63] |
Sun CA, Wu MH, Chu CH, et al. (2010) Adipocytokine resistin and breast cancer risk. Breast Cancer Res Treat 123: 869-876. doi: 10.1007/s10549-010-0792-4
|
| [64] | Dalamaga M, Sotiropoulos G, Karmaniolas K, et al. (2013) Serum resistin: a biomarker of breast cancer in postmenopausal women? Association with clinicopathological characteristics, tumor markers, inflammatory and metabolic parameters. Clin Biochem 46: 584-590. |
| [65] |
Georgiou GP, Provatopoulou X, Kalogera E, et al. (2016) Serum resistin is inversely related to breast cancer risk in premenopausal women. Breast 29: 163-169. doi: 10.1016/j.breast.2016.07.025
|
| [66] |
Rahbar AR, Nabipour I (2014) The relationship between dietary lipids and serum visfatin and adiponectin levels in postmenopausal women. Endocr Metab Immune Disord Drug Targets 14: 84-92. doi: 10.2174/1871530314666140527143009
|
| [67] |
Dalamaga M, Karmaniolas K, Papadavid E, et al. (2011) Elevated serum visfatin/nicotinamide phosphoribosyl-transferase levels are associated with risk of postmenopausal breast cancer independently from adiponectin, leptin, and anthropometric and metabolic parameters. Menopause 18: 1198-1204. doi: 10.1097/gme.0b013e31821e21f5
|
| [68] |
Lee YC, Yang YH, Su JH, et al. (2011) High visfatin expression in breast cancer tissue is associated with poor survival. Cancer Epidemiol Biomarkers Prev 20: 1892-1901. doi: 10.1158/1055-9965.EPI-11-0399
|
| [69] |
Kim SR, Park HJ, Bae YH, et al. (2012) Curcumin down-regulates visfatin expression and inhibits breast cancer cell invasion. Endocrinology 153: 554-563. doi: 10.1210/en.2011-1413
|
| [70] |
Li C, Li J, Chen Y, et al. (2016) Effect of curcumin on visfatin and zinc-alpha2-glycoprotein in a rat model of non-alcoholic fatty liver disease. Acta Cir Bras 31: 706-713. doi: 10.1590/s0102-865020160110000001
|
| [71] |
Gao Y, Wang C, Pan T, et al. (2014) Impact of metformin treatment and swimming exercise on visfatin levels in high-fat-induced obesity rats. Arq Bras Endocrinol Metabol 58: 42-47. doi: 10.1590/0004-2730000002840
|
| [72] |
Maffei M, Halaas J, Ravussin E, et al. (1995) Leptin levels in human and rodent: measurement of plasma leptin and ob RNA in obese and weight-reduced subjects. Nat Med 1: 1155-1161. doi: 10.1038/nm1195-1155
|
| [73] |
Thompson HJ, Sedlacek SM, Wolfe P, et al. (2015) Impact of Weight Loss on Plasma Leptin and Adiponectin in Overweight-to-Obese Post Menopausal Breast Cancer Survivors. Nutrients 7: 5156-5176. doi: 10.3390/nu7075156
|
| [74] |
Alshaker H, Wang Q, Frampton AE, et al. (2015) Sphingosine kinase 1 contributes to leptin-induced STAT3 phosphorylation through IL-6/gp130 transactivation in oestrogen receptor-negative breast cancer. Breast Cancer Res Treat 149: 59-67. doi: 10.1007/s10549-014-3228-8
|
| [75] |
Park HS, Park JY, Yu R (2005) Relationship of obesity and visceral adiposity with serum concentrations of CRP, TNF-alpha and IL-6. Diabetes Res Clin Pract 69: 29-35. doi: 10.1016/j.diabres.2004.11.007
|
| [76] |
Bowers LW, Brenner AJ, Hursting SD, et al. (2015) Obesity-associated systemic interleukin-6 promotes pre-adipocyte aromatase expression via increased breast cancer cell prostaglandin E2 production. Breast Cancer Res Treat 149: 49-57. doi: 10.1007/s10549-014-3223-0
|
| [77] |
Imayama I, Ulrich CM, Alfano CM, et al. (2012) Effects of a caloric restriction weight loss diet and exercise on inflammatory biomarkers in overweight/obese postmenopausal women: a randomized controlled trial. Cancer Res 72: 2314-2326. doi: 10.1158/0008-5472.CAN-11-3092
|
| [78] | Gao D, Rahbar R, Fish EN (2016) CCL5 activation of CCR5 regulates cell metabolism to enhance proliferation of breast cancer cells. Open Biol 6. |
| [79] |
Svensson S, Abrahamsson A, Rodriguez GV, et al. (2015) CCL2 and CCL5 Are Novel Therapeutic Targets for Estrogen-Dependent Breast Cancer. Clin Cancer Res 21: 3794-3805. doi: 10.1158/1078-0432.CCR-15-0204
|
| [80] |
Yaal-Hahoshen N, Shina S, Leider-Trejo L, et al. (2006) The chemokine CCL5 as a potential prognostic factor predicting disease progression in stage II breast cancer patients. Clin Cancer Res 12: 4474-4480. doi: 10.1158/1078-0432.CCR-06-0074
|
| [81] | D'Esposito V, Liguoro D, Ambrosio MR, et al. (2016) Adipose microenvironment promotes triple negative breast cancer cell invasiveness and dissemination by producing CCL5. Oncotarget 7: 24495-24509. |
| [82] |
Li BH, He FP, Yang X, et al. (2017) Steatosis induced CCL5 contributes to early-stage liver fibrosis in nonalcoholic fatty liver disease progress. Transl Res 180: 103-117 e104. doi: 10.1016/j.trsl.2016.08.006
|
| [83] |
Bell LN, Ward JL, Degawa-Yamauchi M, et al. (2006) Adipose tissue production of hepatocyte growth factor contributes to elevated serum HGF in obesity. Am J Physiol Endocrinol Metab 291: E843-848. doi: 10.1152/ajpendo.00174.2006
|
| [84] |
Sundaram S, Freemerman AJ, Johnson AR, et al. (2013) Role of HGF in obesity-associated tumorigenesis: C3(1)-TAg mice as a model for human basal-like breast cancer. Breast Cancer Res Treat 142: 489-503. doi: 10.1007/s10549-013-2741-5
|
| [85] | Ristimaki A, Sivula A, Lundin J, et al. (2002) Prognostic significance of elevated cyclooxygenase-2 expression in breast cancer. Cancer Res 62: 632-635. |
| [86] |
Khuder SA, Mutgi AB (2001) Breast cancer and NSAID use: a meta-analysis. Br J Cancer 84: 1188-1192. doi: 10.1054/bjoc.2000.1709
|
| [87] | Brown KA, Simpson ER (2012) Obesity and breast cancer: mechanisms and therapeutic implications. Front Biosci (Elite Ed) 4: 2515-2524. |
| [88] | Davies NJ, Batehup L, Thomas R (2011) The role of diet and physical activity in breast, colorectal, and prostate cancer survivorship: a review of the literature. Br J Cancer 105 Suppl 1: S52-73. |
| [89] |
Cuzick J, Warwick J, Pinney E, et al. (2011) Tamoxifen-induced reduction in mammographic density and breast cancer risk reduction: a nested case-control study. J Natl Cancer Inst 103: 744-752. doi: 10.1093/jnci/djr079
|
| [90] |
Kaaks R, Bellati C, Venturelli E, et al. (2003) Effects of dietary intervention on IGF-I and IGF-binding proteins, and related alterations in sex steroid metabolism: the Diet and Androgens (DIANA) Randomised Trial. Eur J Clin Nutr 57: 1079-1088. doi: 10.1038/sj.ejcn.1601647
|
| [91] | Wei M, Brandhorst S, Shelehchi M, et al. (2017) Fasting-mimicking diet and markers/risk factors for aging, diabetes, cancer, and cardiovascular disease. Sci Transl Med 9. |
| [92] |
Sandhu N, Schetter SE, Liao J, et al. (2016) Influence of Obesity on Breast Density Reduction by Omega-3 Fatty Acids: Evidence from a Randomized Clinical Trial. Cancer Prev Res (Phila) 9: 275-282. doi: 10.1158/1940-6207.CAPR-15-0235
|
| [93] |
Sanderson M, Peltz G, Perez A, et al. (2010) Diabetes, physical activity and breast cancer among Hispanic women. Cancer Epidemiol 34: 556-561. doi: 10.1016/j.canep.2010.06.001
|
| [94] |
Irwin ML, Varma K, Alvarez-Reeves M, et al. (2009) Randomized controlled trial of aerobic exercise on insulin and insulin-like growth factors in breast cancer survivors: the Yale Exercise and Survivorship study. Cancer Epidemiol Biomarkers Prev 18: 306-313. doi: 10.1158/1055-9965.EPI-08-0531
|
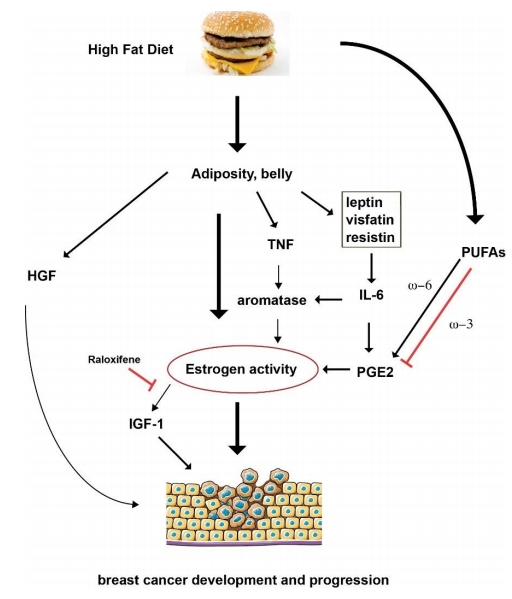
Figures(1)

Xin Nian, Yasuhiro Nagai, Cameron Jeffers, Kara N. Maxwell, Hongtao Zhang. Dietary influence on estrogens and cytokines in breast cancer[J]. AIMS Molecular Science, 2017, 4(3): 252-270. doi: 10.3934/molsci.2017.3.252







 DownLoad:
DownLoad: