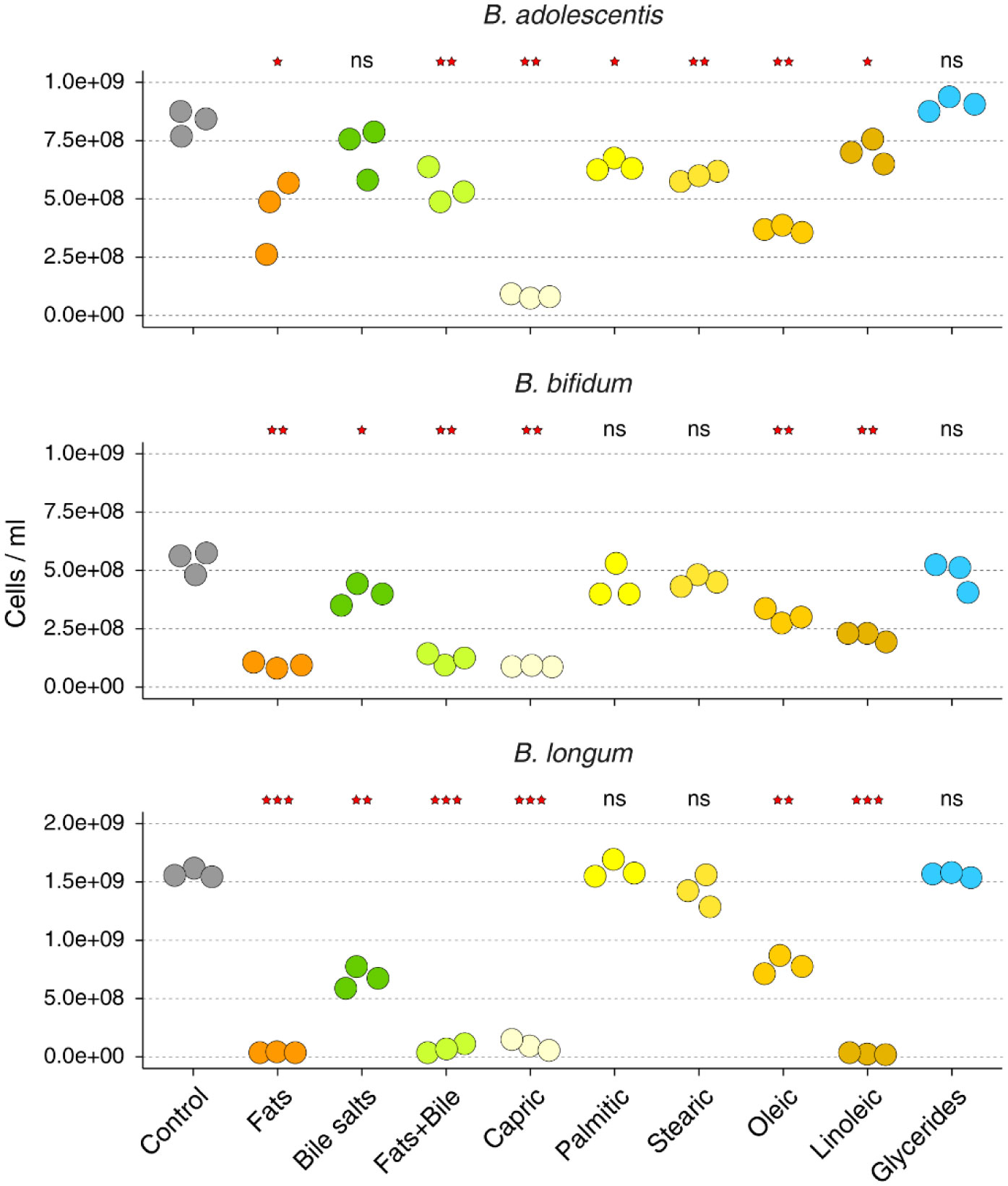
High-fat diets have been associated with lower gut and fecal abundances of genus Bifidobacterium. Here, we investigated whether commonly consumed dietary free fatty acids have any detrimental effect on the growth of B. adolescentis, B. bifidum, and B. longum. We found that the presence of free fatty acids in the medium inhibits the growth of Bifidobacterium species to a varying degree, with capric (C10:0), oleic (C18:1), and linoleic (C18:2) acids displaying the largest effect. In comparison, free fatty acids did not affect the growth of Escherichia coli. When fats were added as a mixture of mono- and diacylglycerols, the inhibitory effect on Bifidobacterium growth was abolished.
Citation: Sergio Perez-Burillo, Sumudu Rajakaruna, Oleg Paliy. Growth of Bifidobacterium species is inhibited by free fatty acids and bile salts but not by glycerides[J]. AIMS Microbiology, 2022, 8(1): 53-60. doi: 10.3934/microbiol.2022005
High-fat diets have been associated with lower gut and fecal abundances of genus Bifidobacterium. Here, we investigated whether commonly consumed dietary free fatty acids have any detrimental effect on the growth of B. adolescentis, B. bifidum, and B. longum. We found that the presence of free fatty acids in the medium inhibits the growth of Bifidobacterium species to a varying degree, with capric (C10:0), oleic (C18:1), and linoleic (C18:2) acids displaying the largest effect. In comparison, free fatty acids did not affect the growth of Escherichia coli. When fats were added as a mixture of mono- and diacylglycerols, the inhibitory effect on Bifidobacterium growth was abolished.

| [1] |
Trumbo P, Schlicker S, Yates AA, et al. (2002) Dietary reference intakes for energy, carbohydrate, fiber, fat, fatty acids, cholesterol, protein and amino acids. J Am Diet Assoc 102: 1621-1630. https://doi.org/10.1016/S0002-8223(02)90346-9 
|
| [2] | Wang TY, Liu M, Portincasa P, et al. (2013) New insights into the molecular mechanism of intestinal fatty acid absorption. Eur J Clin Invest 43: 1203-1223. https://doi.org/10.1111/eci.12161 |
| [3] |
Mu H, Hoy CE (2004) The digestion of dietary triacylglycerols. Prog Lipid Res 43: 105-133. https://doi.org/10.1016/S0163-7827(03)00050-X
|
| [4] |
Iqbal J, Hussain MM (2009) Intestinal lipid absorption. Am J Physiol Endocrinol Metab 296: E1183-1194. https://doi.org/10.1152/ajpendo.90899.2008
|
| [5] | Hoyles L, Wallace RJ (2010) Gastrointestinal Tract: Fat Metabolism in the Colon. Handbook of Hydrocarbon and Lipid Microbiology . Berlin: Heidelberg: Springer. https://doi.org/10.1007/978-3-540-77587-4_233 |
| [6] |
Shortt C, Hasselwander O, Meynier A, et al. (2018) Systematic review of the effects of the intestinal microbiota on selected nutrients and non-nutrients. Eur J Nutr 57: 25-49. https://doi.org/10.1007/s00394-017-1546-4
|
| [7] |
Agans R, Gordon A, Kramer DL, et al. (2018) Dietary Fatty Acids Sustain the Growth of the Human Gut Microbiota. Appl Environ Microbiol 84: e01525-01518. https://doi.org/10.1128/AEM.01525-18
|
| [8] |
Lindefeldt M, Eng A, Darban H, et al. (2019) The ketogenic diet influences taxonomic and functional composition of the gut microbiota in children with severe epilepsy. NPJ Biofilms Microbiomes 5: 5. https://doi.org/10.1038/s41522-018-0073-2
|
| [9] |
Brinkworth GD, Noakes M, Clifton PM, et al. (2009) Comparative effects of very low-carbohydrate, high-fat and high-carbohydrate, low-fat weight-loss diets on bowel habit and faecal short-chain fatty acids and bacterial populations. Br J Nutr 101: 1493-1502. https://doi.org/10.1017/S0007114508094658
|
| [10] |
Cani PD, Neyrinck AM, Fava F, et al. (2007) Selective increases of bifidobacteria in gut microflora improve high-fat-diet-induced diabetes in mice through a mechanism associated with endotoxaemia. Diabetologia 50: 2374-2383. https://doi.org/10.1007/s00125-007-0791-0
|
| [11] |
Gunasekera TS, Csonka LN, Paliy O (2008) Genome-wide transcriptional responses of Escherichia coli k-12 to continuous osmotic and heat stresses. J Bacteriol 190: 3712-3720. https://doi.org/10.1128/JB.01990-07
|
| [12] |
Zheng CJ, Yoo JS, Lee TG, et al. (2005) Fatty acid synthesis is a target for antibacterial activity of unsaturated fatty acids. FEBS Lett 579: 5157-5162. https://doi.org/10.1016/j.febslet.2005.08.028
|
| [13] |
Agans RT, Gordon A, Hussain S, et al. (2019) Titanium Dioxide Nanoparticles Elicit Lower Direct Inhibitory Effect on Human Gut Microbiota Than Silver Nanoparticles. Toxicol Sci 172: 411-416. https://doi.org/10.1093/toxsci/kfz183
|
| [14] | Benjamini Y, Hochberg Y (1995) Controlling the False Discovery Rate-a practical and powerful approach to multiple testing. J Roy Stat Soc B Met 57: 289-300. https://doi.org/10.1111/j.2517-6161.1995.tb02031.x |
| [15] |
Senizza A, Rocchetti G, Callegari ML, et al. (2020) Linoleic acid induces metabolic stress in the intestinal microorganism Bifidobacterium breve DSM 20213. Sci Rep 10: 5997. https://doi.org/10.1038/s41598-020-62897-w
|
| [16] | Yoon BK, Jackman JA, Valle-González ER, et al. (2018) Antibacterial Free Fatty Acids and Monoglycerides: Biological Activities, Experimental Testing, and Therapeutic Applications. Int J Mol Sci 19. https://doi.org/10.3390/ijms19041114 |
| [17] |
Hovorková P, Laloučková K, Skřivanová E (2018) Determination of in vitro antibacterial activity of plant oils containing medium-chain fatty acids against Gram-positive pathogenic and gut commensal bacteria. Czech J Anim Sci 63: 119-125. https://doi.org/10.17221/70/2017-CJAS
|
| [18] |
Thormar H, Hilmarsson H, Bergsson G (2006) Stable concentrated emulsions of the 1-monoglyceride of capric acid (monocaprin) with microbicidal activities against the food-borne bacteria Campylobacter jejuni, Salmonella spp., and Escherichia coli. Appl Environ Microbiol 72: 522-526. https://doi.org/10.1128/AEM.72.1.522-526.2006
|
| [19] |
Li N, Yan F, Wang N, et al. (2020) Distinct Gut Microbiota and Metabolite Profiles Induced by Different Feeding Methods in Healthy Chinese Infants. Front Microbiol 11: 714. https://doi.org/10.3389/fmicb.2020.00714
|
| [20] |
Ruiz L, Margolles A, Sánchez B (2013) Bile resistance mechanisms in Lactobacillus and Bifidobacterium. Front Microbiol 4: 396. https://doi.org/10.3389/fmicb.2013.00396
|
| [21] |
Hildebrand P, Beglinger C, Gyr K, et al. (1990) Effects of a cholecystokinin receptor antagonist on intestinal phase of pancreatic and biliary responses in man. J Clin Invest 85: 640-646. https://doi.org/10.1172/JCI114486
|
| [22] |
Kim GB, Yi SH, Lee BH (2004) Purification and characterization of three different types of bile salt hydrolases from Bifidobacterium strains. J Dairy Sci 87: 258-266. https://doi.org/10.3168/jds.S0022-0302(04)73164-1
|
| [23] |
Sheu CW, Freese E (1973) Lipopolysaccharide layer protection of gram-negative bacteria against inhibition by long-chain fatty acids. J Bacteriol 115: 869-875. https://doi.org/10.1128/jb.115.3.869-875.1973
|
 microbiol-08-01-005-s001.pdf microbiol-08-01-005-s001.pdf |
 |
Figures(1) / Tables(2)

Sergio Perez-Burillo, Sumudu Rajakaruna, Oleg Paliy. Growth of Bifidobacterium species is inhibited by free fatty acids and bile salts but not by glycerides[J]. AIMS Microbiology, 2022, 8(1): 53-60. doi: 10.3934/microbiol.2022005







 DownLoad:
DownLoad: