Citation: Tru Tran, Stephanie N. Dawrs, Grant J. Norton, Ravleen Virdi, Jennifer R. Honda. Brought to you courtesy of the red, white, and blue–pigments of nontuberculous mycobacteria[J]. AIMS Microbiology, 2020, 6(4): 434-450. doi: 10.3934/microbiol.2020026

| [1] |
Tuli HS, Chaudhary P, Beniwal V, et al. (2015) Microbial pigments as natural color sources: current trends and future perspectives. J Food Sci Technol 52: 4669-4678. doi: 10.1007/s13197-014-1601-6

|
| [2] |
Ramesh C, Vinithkumar NV, Kirubagaran R, et al. (2019) Multifaceted applications of microbial pigments: current knowledge, challenges and future directions for public health implications. Microorganisms 7. doi: 10.3390/microorganisms7070186
|
| [3] | Kothari V, Joshi C, Patel P (2016) Colourful side of bacteriology: the pigmented bacteria. Adv Genet Eng 5. |
| [4] |
Valeru SP, Rompikuntal PK, Ishikawa T, et al. (2009) Role of melanin pigment in expression of Vibrio cholerae virulence factors. Infect Immun 77: 935-942. doi: 10.1128/IAI.00929-08
|
| [5] |
Gonzalez JE, Keshavan ND (2006) Messing with bacterial quorum sensing. Microbiol Mol Biol Rev 70: 859-875. doi: 10.1128/MMBR.00002-06
|
| [6] |
Devescovi G, Kojic M, Covaceuszach S, et al. (2017) Negative regulation of violacein biosynthesis in Chromobacterium violaceum. Front Microbiol 8: 349. doi: 10.3389/fmicb.2017.00349
|
| [7] |
Baker JA (1938) Light as a factor in the production of pigment by certain bacteria. J Bacteriol 35: 625-631. doi: 10.1128/JB.35.6.625-631.1938
|
| [8] |
Lin YB, Wang XY, Fang H, et al. (2012) Streptomyces shaanxiensis sp. nov., a blue pigment-producing streptomycete from sewage irrigation soil. Int J Syst Evol Microbiol 62: 1725-1730. doi: 10.1099/ijs.0.029959-0
|
| [9] |
Kirti K, Amita S, Priti S, et al. (2014) Colorful world of microbes: carotenoids and their applications. Adv Biol 2014: 1-13. doi: 10.1155/2014/837891
|
| [10] | Azmana AS, Mawangb CI, Abubakara S (2018) Bacterial pigments: the bioactivities and as an alternative for therapeutic applications. Nat Prod Commun 13: 1747-1754. |
| [11] |
Nosanchuk JD, Casadevall A (2006) Impact of melanin on microbial virulence and clinical resistance to antimicrobial compounds. Antimicrob Agents Chemother 50: 3519-3528. doi: 10.1128/AAC.00545-06
|
| [12] |
Turick CE, Beliaev AS, Zakrajsek BA, et al. (2009) The role of 4-hydroxyphenylpyruvate dioxygenase in enhancement of solid-phase electron transfer by Shewanella oneidensis MR-1. FEMS Microbiol Ecol 68: 223-225. doi: 10.1111/j.1574-6941.2009.00670.x
|
| [13] |
Zheng H, Chatfield CH, Liles MR, et al. (2013) Secreted pyomelanin of Legionella pneumophila promotes bacterial iron uptake and growth under iron-limiting conditions. Infect Immun 81: 4182-4191. doi: 10.1128/IAI.00858-13
|
| [14] |
Pavan ME, Lopez NI, Pettinari MJ (2020) Melanin biosynthesis in bacteria, regulation and production perspectives. Appl Microbiol Biotechnol 104: 1357-1370. doi: 10.1007/s00253-019-10245-y
|
| [15] |
Castro-Sowinski S, Matan O, Bonafede P, et al. (2007) A thioredoxin of Sinorhizobium meliloti CE52G is required for melanin production and symbiotic nitrogen fixation. Mol Plant Microbe Interact 20: 986-993. doi: 10.1094/MPMI-20-8-0986
|
| [16] |
Yang D, Kim WJ, Yoo SM, et al. (2018) Repurposing type III polyketide synthase as a malonyl-CoA biosensor for metabolic engineering in bacteria. Proc Natl Acad Sci USA 115: 9835-9844. doi: 10.1073/pnas.1808567115
|
| [17] |
Ikeda K, Masujima T, Sugiyama M (1996) Effects of methionine and Cu2+ on the expression of tyrosinase activity in Streptomyces castaneoglobisporus. J Biochem 120: 1141-1145. doi: 10.1093/oxfordjournals.jbchem.a021533
|
| [18] |
Zhu D, He X, Zhou X, et al. (2005) Expression of the melC operon in several Streptomyces strains is positively regulated by AdpA, an AraC family transcriptional regulator involved in morphological development in Streptomyces coelicolor. J Bacteriol 187: 3180-3187. doi: 10.1128/JB.187.9.3180-3187.2005
|
| [19] |
Zughaier SM, Ryley HC, Jackson SK (1999) A melanin pigment purified from an epidemic strain of Burkholderia cepacia attenuates monocyte respiratory burst activity by scavenging superoxide anion. Infect Immun 67: 908-913. doi: 10.1128/IAI.67.2.908-913.1999
|
| [20] |
Keith KE, Killip L, He P, et al. (2007) Burkholderia cenocepacia C5424 produces a pigment with antioxidant properties using a homogentisate intermediate. J Bacteriol 189: 9057-9065. doi: 10.1128/JB.00436-07
|
| [21] |
Noorian P, Hu J, Chen Z, et al. (2017) Pyomelanin produced by Vibrio cholerae confers resistance to predation by Acanthamoeba castellanii. FEMS Microbiol Ecol 93. doi: 10.1093/femsec/fix147
|
| [22] |
El-Fouly MZ SA, Shahin AAM, El-Bialy HA, et al. (2015) Biosynthesis of pyocyanin pigment by Pseudomonas aeruginosa. J Radiat Res Appl Sci 8: 36-48. doi: 10.1016/j.jrras.2014.10.007
|
| [23] |
Jayaseelan S, Ramaswamy D, Dharmaraj S (2014) Pyocyanin: production, applications, challenges and new insights. World J Microbiol Biotechnol 30: 1159-1168. doi: 10.1007/s11274-013-1552-5
|
| [24] |
Price-Whelan A, Dietrich LE, Newman DK (2007) Pyocyanin alters redox homeostasis and carbon flux through central metabolic pathways in Pseudomonas aeruginosa PA14. J Bacteriol 189: 6372-6381. doi: 10.1128/JB.00505-07
|
| [25] |
Lau GW, Ran H, Kong F, et al. (2004) Pseudomonas aeruginosa pyocyanin is critical for lung infection in mice. Infect Immun 72: 4275-4278. doi: 10.1128/IAI.72.7.4275-4278.2004
|
| [26] |
Saha S, Thavasi R, Jayalakshmi S (2008) Phenazine pigments from Pseudomonas aeruginosa and their application as antibacterial agent and food colourants. Res J Microbiol 3: 122-128. doi: 10.3923/jm.2008.122.128
|
| [27] |
Satapute PK, Kaliwal B (2016) Biodegradation of the fungicide propiconazole by Pseudomonas aeruginosa PS-4 strain isolated from a paddy soil. Ann Microbiol 66: 1355-1365. doi: 10.1007/s13213-016-1222-6
|
| [28] |
Kavitha K, Mathiyazhagan S, Sendhilvel V, et al. (2007) Broad spectrum action of phenazine against active and dormant structures of fungal pathogens and root knot nematode. Arch Phytopathol Plant Prot 38: 69-76. doi: 10.1080/03235400400008408
|
| [29] |
Winstanley C, Fothergill JL (2009) The role of quorum sensing in chronic cystic fibrosis Pseudomonas aeruginosa infections. FEMS Microbiol Lett 290: 1-9. doi: 10.1111/j.1574-6968.2008.01394.x
|
| [30] |
Mavrodi DV, Bonsall RF, Delaney SM, et al. (2001) Functional analysis of genes for biosynthesis of pyocyanin and phenazine-1-carboxamide from Pseudomonas aeruginosa PAO1. J Bacteriol 183: 6454-6465. doi: 10.1128/JB.183.21.6454-6465.2001
|
| [31] |
Greenhagen BT, Shi K, Robinson H, et al. (2008) Crystal structure of the pyocyanin biosynthetic protein PhzS. Biochemistry 47: 5281-5289. doi: 10.1021/bi702480t
|
| [32] |
DeBritto S, Gajbar TD, Satapute P, et al. (2020) Isolation and characterization of nutrient dependent pyocyanin from Pseudomonas aeruginosa and its dye and agrochemical properties. Sci Rep 10: 1542. doi: 10.1038/s41598-020-58335-6
|
| [33] | Devnath P, Uddin K, Ahamed F, et al. (2017) Extraction, purification and characterization of pyocyanin produced by Pseudomonas aeruginosa and evaluation for its antimicrobial activity. Int Res J Biol Sci 6: 1-9. |
| [34] |
Castaneda-Tamez P, Ramirez-Peris J, Perez-Velazquez J, et al. (2018) Pyocyanin restricts social cheating in Pseudomonas aeruginosa. Front Microbiol 9: 1348. doi: 10.3389/fmicb.2018.01348
|
| [35] |
Price-Whelan A, Dietrich LE, Newman DK (2006) Rethinking ‘secondary’ metabolism: physiological roles for phenazine antibiotics. Nat Chem Biol 2: 71-78. doi: 10.1038/nchembio764
|
| [36] |
Allen L, Dockrell DH, Pattery T, et al. (2005) Pyocyanin production by Pseudomonas aeruginosa induces neutrophil apoptosis and impairs neutrophil-mediated host defenses in vivo. J Immunol 174: 3643-3649. doi: 10.4049/jimmunol.174.6.3643
|
| [37] |
Hiraishi A, Shimada K (2001) Aerobic anoxygenic photosynthetic bacteria with zinc-bacteriochlorophyll. J Gen Appl Microbiol 47: 161-180. doi: 10.2323/jgam.47.161
|
| [38] | Willows RD, Kriegel AM (2009) The Purple Phototrophic Bacteria. Advances in Photosynthesis and Respiration Dordrecht: Springer. |
| [39] |
Harada J, Mizoguchi T, Tsukatani Y, et al. (2012) A seventh bacterial chlorophyll driving a large light-harvesting antenna. Sci Rep 2: 671. doi: 10.1038/srep00671
|
| [40] |
Brandis AS, Salomon Y, Scherz A (2006) Bacteriochlorophyll Sensitizers in Photodynamic Therapy. Chlorophylls and Bacteriochlorophylls Advances in Photosynthesis and Respiration Dordrecht: Springer, 485-494. doi: 10.1007/1-4020-4516-6_33
|
| [41] |
Limantara L, Koehler P, Wilhelm B, et al. (2006) Photostability of bacteriochlorophyll a and derivatives: potential sensitizers for photodynamic tumor therapy. Photochem Photobiol 82: 770-780. doi: 10.1562/2005-09-07-RA-676
|
| [42] |
Blankenship RE (2004) Identification of a key step in the biosynthetic pathway of bacteriochlorophyll c and its implications for other known and unknown green sulfur bacteria. J Bacteriol 186: 5187-5188. doi: 10.1128/JB.186.16.5187-5188.2004
|
| [43] | Bryant N-UFGMCAMA (2006) Chlorophylls and Bacteriochlorophylls. Advances in Photosynthesis and Respiration Dordrecht: Springer. |
| [44] | Choi SY, Yoon KH, Lee JI, et al. (2014) Violacein: properties and production of a versatile bacterial pigment. BioMed Res Int 2015: 1-8. |
| [45] |
de Siqueira IC, Dias J, Ruf H, et al. (2005) Chromobacterium violaceum in siblings, Brazil. Emerg Infect Dis 11: 1443-1445. doi: 10.3201/eid1109.050278
|
| [46] |
Konzen M, De Marco D, Cordova CA, et al. (2006) Antioxidant properties of violacein: possible relation on its biological function. Bioorg Med Chem 14: 8307-8313. doi: 10.1016/j.bmc.2006.09.013
|
| [47] |
Ferreira CV, Bos CL, Versteeg HH, et al. (2004) Molecular mechanism of violacein-mediated human leukemia cell death. Blood 104: 1459-1464. doi: 10.1182/blood-2004-02-0594
|
| [48] |
Matz C, Deines P, Boenigk J, et al. (2004) Impact of violacein-producing bacteria on survival and feeding of bacterivorous nanoflagellates. Appl Environ Microbiol 70: 1593-1599. doi: 10.1128/AEM.70.3.1593-1599.2004
|
| [49] | Choi SY, Yoon KH, Lee JI, et al. (2015) Violacein: properties and production of a versatile bacterial pigment. Biomed Res Int 2015: 465056. |
| [50] |
Fuller JJ, Ropke R, Krausze J, et al. (2016) Biosynthesis of violacein, structure and function of l-Tryptophan oxidase VioA from Chromobacterium violaceum. J Biol Chem 291: 20068-20084. doi: 10.1074/jbc.M116.741561
|
| [51] |
Nakamura Y, Asada C, Sawada T (2003) Production of antibacterial violet pigment by psychrotropic bacterium RT102 strain. Biotechnol Bioprocess Eng 8: 37-40. doi: 10.1007/BF02932896
|
| [52] |
Subramaniam S, Ravi V, Sivasubramanian A (2014) Synergistic antimicrobial profiling of violacein with commercial antibiotics against pathogenic micro-organisms. Pharm Biol 52: 86-90. doi: 10.3109/13880209.2013.815634
|
| [53] |
Melo PS, Maria SS, Vidal BC, et al. (2000) Violacein cytotoxicity and induction of apoptosis in V79 cells. In Vitro Cell Dev Biol Anim 36: 539-543. doi: 10.1290/1071-2690(2000)036<0539:VCAIOA>2.0.CO;2
|
| [54] |
de Carvalho DD, Costa FT, Duran N, et al. (2006) Cytotoxic activity of violacein in human colon cancer cells. Toxicol In Vitro 20: 1514-1521. doi: 10.1016/j.tiv.2006.06.007
|
| [55] |
Bilsland E, Tavella TA, Krogh R, et al. (2018) Antiplasmodial and trypanocidal activity of violacein and deoxyviolacein produced from synthetic operons. BMC Biotechnol 18: 22. doi: 10.1186/s12896-018-0428-z
|
| [56] |
Sakai-Kawada FE, Ip CG, Hagiwara KA, et al. (2019) Biosynthesis and bioactivity of prodiginine analogs in marine bacteria, pseudoalteromonas: a mini review. Front Microbiol 10: 1715. doi: 10.3389/fmicb.2019.01715
|
| [57] |
Boric M, Danevcic T, Stopar D (2011) Prodigiosin from Vibrio sp. DSM 14379; a new UV-protective pigment. Microb Ecol 62: 528-536. doi: 10.1007/s00248-011-9857-0
|
| [58] | Nwankwo IU IV, Chidiebere OL, Nwachukwu MP (2017) Evaluation of antimicrobial activity of prodigiosin produced from Serratia marcescens against some pathogenic bacteria. Futo J Series 3: 93-102. |
| [59] | Singh P, Shekhawat N (2012) Chemometric descriptors in the rationale of antimalarial activity of natural and synthetic prodiginines. J Curr Chem Pharm Sci 2: 224-260. |
| [60] |
Suryawanshi RK, Koujah L, Patil CD, et al. (2020) Bacterial pigment prodigiosin demonstrates a unique antiherpesvirus activity that is mediated through inhibition of prosurvival signal transducers. J Virol 94. doi: 10.1128/JVI.00251-20
|
| [61] |
Darshan N, Manonmani HK (2015) Prodigiosin and its potential applications. J Food Sci Technol 52: 5393-5407. doi: 10.1007/s13197-015-1740-4
|
| [62] |
Gristwood T, McNeil MB, Clulow JS, et al. (2011) PigS and PigP regulate prodigiosin biosynthesis in Serratia via differential control of divergent operons, which include predicted transporters of sulfur-containing molecules. J Bacteriol 193: 1076-1085. doi: 10.1128/JB.00352-10
|
| [63] | Pandey R, Chander R, Sainis KB (2007) Prodigiosins: a novel family of immunosuppressants with anti-cancer activity. Indian J Biochem Biophys 44: 295-302. |
| [64] |
Montaner B, Perez-Tomas R (2001) Prodigiosin-induced apoptosis in human colon cancer cells. Life Sci 68: 2025-2036. doi: 10.1016/S0024-3205(01)01002-5
|
| [65] | Kimyon O, Das T, Ibugo AI, et al. (2016) Serratia secondary metabolite prodigiosin inhibits Pseudomonas aeruginosa biofilm development by producing reactive oxygen species that damage biological molecules. Front Microbiol 7: 972. |
| [66] |
Van Dien SJ, Marx CJ, O'Brien BN, et al. (2003) Genetic characterization of the carotenoid biosynthetic pathway in Methylobacterium extorquens AM1 and isolation of a colorless mutant. Appl Environ Microbiol 69: 7563-7566. doi: 10.1128/AEM.69.12.7563-7566.2003
|
| [67] |
Frengova GI, Beshkova DM (2009) Carotenoids from Rhodotorula and Phaffia: yeasts of biotechnological importance. J Ind Microbiol Biotechnol 36: 163-180. doi: 10.1007/s10295-008-0492-9
|
| [68] |
Cooney JJ, Marks HW, Smith AM (1966) Isolation and Identification of Canthaxanthin from Micrococcus roseus. J Bacteriol 92: 342-345. doi: 10.1128/JB.92.2.342-345.1966
|
| [69] |
Miller NJ, Sampson J, Candeias LP, et al. (1996) Antioxidant activities of carotenes and xanthophylls. FEBS Lett 384: 240-242. doi: 10.1016/0014-5793(96)00323-7
|
| [70] |
Jagannadham MV, Chattopadhyay MK, Subbalakshmi C, et al. (2000) Carotenoids of an Antarctic psychrotolerant bacterium, Sphingobacterium antarcticus, and a mesophilic bacterium, Sphingobacterium multivorum. Arch Microbiol 173: 418-424. doi: 10.1007/s002030000163
|
| [71] |
Liu GY, Essex A, Buchanan JT, et al. (2005) Staphylococcus aureus golden pigment impairs neutrophil killing and promotes virulence through its antioxidant activity. J Exp Med 202: 209-215. doi: 10.1084/jem.20050846
|
| [72] |
Rajagopal L, Sundari CS, Balasubramanian D, et al. (1997) The bacterial pigment xanthomonadin offers protection against photodamage. FEBS Lett 415: 125-128. doi: 10.1016/S0014-5793(97)01109-5
|
| [73] |
Dieser M, Greenwood M, Foreman CM (2010) Carotenoid pigmentation in Antarctic hetero-trophic bacteria as a strategy to withstand environmental stresses. Arct Antarct Alp Res 42: 396-405. doi: 10.1657/1938-4246-42.4.396
|
| [74] | Koyyati R, Kudle KR, Padigya PRM (2019) Antibacterial, antioxidant and cytotoxic activity of bacterial carotenoids isolated from Rhodopseudomonas palustris KRPR01 and KRPR02. Int J Pharm Sci Res 10: 4644-4649. |
| [75] | Ravikumar S UG, Gokulakrishnan R (2016) Antibacterial property of Halobacterial carotenoids against human bacterial pathogens. J Sci Ind (India) 75: 253-257. |
| [76] |
Paniagua-Michel J, Olmos-Soto J, Ruiz MA (2012) Pathways of carotenoid biosynthesis in bacteria and microalgae. Methods Mol Biol 892: 1-12. doi: 10.1007/978-1-61779-879-5_1
|
| [77] |
Chew BP, Park JS (2004) Carotenoid action on the immune response. J Nutr 134: 257S-261S. doi: 10.1093/jn/134.1.257S
|
| [78] | Azman AS, Mawang CI, Abubakar S (2018) Bacterial Pigments: The Bioactivities and as an Alternative for Therapeutic Applications. Nat Prod Commun 13: 1747-1754. |
| [79] |
Reichenbach H, Kleinig H, Achenbach H (1974) The pigments of Flexibacter elegans: Novel and chemosystematically useful compounds. Arch Microbiol 101: 131-144. doi: 10.1007/BF00455933
|
| [80] |
Ghosh A, Goyal A, Jain RK (2007) Study of methanol-induced phenotypic changes in a novel strain of Acinetobacter lwoffii. Arch Microbiol 188: 533-539. doi: 10.1007/s00203-007-0268-z
|
| [81] | Margalith PZ (1992) Pigment Microbiology. Pigment Microbiology London, UK: Chapman & Hall. |
| [82] |
Ivanov AG, Krol M, Selstam E, et al. (2007) The induction of CP43' by iron-stress in Synechococcus sp. PCC 7942 is associated with carotenoid accumulation and enhanced fatty acid unsaturation. Biochim Biophys Acta 1767: 807-813. doi: 10.1016/j.bbabio.2007.02.006
|
| [83] |
Howes CD, Batra PP (1970) Accumulation of lycopene and inhibition of cyclic carotenoids in Mycobacterium in the presence of nicotine. Biochim Biophys Acta 222: 174-179. doi: 10.1016/0304-4165(70)90362-4
|
| [84] |
Saviola B, Felton J (2011) Acidochromogenicity is a common characteristic in nontuberculous mycobacteria. BMC Res Notes 4: 466. doi: 10.1186/1756-0500-4-466
|
| [85] | Saviola B (2014) Pigments and pathogenesis. J Mycobact Dis 4: 1-3. |
| [86] |
Coyne VE, al-Harthi L (1992) Induction of melanin biosynthesis in Vibrio cholerae. Appl Environ Microbiol 58: 2861-2865. doi: 10.1128/AEM.58.9.2861-2865.1992
|
| [87] | Euseby J (1997) List of prokaryotic names with standing in nomenclature. |
| [88] |
Runyon EH (1970) Pigment variations in photochromogenic mycobacteria with special reference to M. vaccae. Pneumonologie 142: 90-93. doi: 10.1007/BF02095201
|
| [89] |
Robledo JA, Murillo AM, Rouzaud F (2011) Physiological role and potential clinical interest of mycobacterial pigments. IUBMB Life 63: 71-78. doi: 10.1002/iub.424
|
| [90] |
Tarnok I, Tarnok Z (1970) Carotene and xanthophylls in mycobacteria. I. Technical procedures; thin-layer chromatographic patterns of mycobacterial pigments. Tubercle 51: 305-312. doi: 10.1016/0041-3879(70)90024-3
|
| [91] |
Ramakrishnan L, Tran HT, Federspiel NA, et al. (1997) A crtB homolog essential for photochromogenicity in Mycobacterium marinum: isolation, characterization, and gene disruption via homologous recombination. J Bacteriol 179: 5862-5868. doi: 10.1128/JB.179.18.5862-5868.1997
|
| [92] |
Provvedi R, Kocincova D, Dona V, et al. (2008) SigF controls carotenoid pigment production and affects transformation efficiency and hydrogen peroxide sensitivity in Mycobacterium smegmatis. J Bacteriol 190: 7859-7863. doi: 10.1128/JB.00714-08
|
| [93] |
Singh AK, Dutta D, Singh V, et al. (2015) Characterization of Mycobacterium smegmatis sigF mutant and its regulon: overexpression of SigF antagonist (MSMEG_1803) in M. smegmatis mimics sigF mutant phenotype, loss of pigmentation, and sensitivity to oxidative stress. Microbiologyopen 4: 896-916. doi: 10.1002/mbo3.288
|
| [94] |
Runyon EH (1959) Anonymous mycobacteria in pulmonary disease. Med Clin North Am 43: 273-290. doi: 10.1016/S0025-7125(16)34193-1
|
| [95] |
Koh WJ, Kwon OJ, Lee KS (2002) Nontuberculous mycobacterial pulmonary diseases in immunocompetent patients. Korean J Radiol 3: 145-157. doi: 10.3348/kjr.2002.3.3.145
|
| [96] |
Kiehn TE, Edwards FF, Brannon P, et al. (1985) Infections caused by Mycobacterium avium complex in immunocompromised patients: diagnosis by blood culture and fecal examination, antimicrobial susceptibility tests, and morphological and seroagglutination characteristics. J Clin Microbiol 21: 168-173. doi: 10.1128/JCM.21.2.168-173.1985
|
| [97] |
van Ingen J, Turenne CY, Tortoli E, et al. (2018) A definition of the Mycobacterium avium complex for taxonomical and clinical purposes, a review. Int J Syst Evol Microbiol 68: 3666-3677. doi: 10.1099/ijsem.0.003026
|
| [98] |
Honda JR, Hasan NA, Davidson RM, et al. (2016) Environmental Nontuberculous Mycobacteria in the Hawaiian Islands. PLoS Negl Trop Dis 10: e0005068. doi: 10.1371/journal.pntd.0005068
|
| [99] |
Falkinham JO, Williams MD, Kwait R, et al. (2016) Methylobacterium spp. as an indicator for the presence or absence of Mycobacterium spp. Int J Mycobacteriol 5: 240-243. doi: 10.1016/j.ijmyco.2016.03.001
|
| [100] |
Garcia-Coca M, Rodriguez-Sevilla G, Perez-Domingo A, et al. (2020) Inhibition of Mycobacterium abscessus, M. chelonae, and M. fortuitum biofilms by Methylobacterium sp. J Antibiot (Tokyo) 73: 40-47. doi: 10.1038/s41429-019-0232-6
|
| [101] |
Frehel C, Ryter A, Rastogi N, et al. (1986) The electron-transparent zone in phagocytized Mycobacterium avium and other mycobacteria: formation, persistence and role in bacterial survival. Ann Inst Pasteur Microbiol 137B: 239-257. doi: 10.1016/S0769-2609(86)80115-6
|
| [102] |
Gordon RE, Pang CH (1970) Black beauty out of Mycobacterium fortuitum Cruz. Appl Microbiol 19: 862-864. doi: 10.1128/AEM.19.5.862-864.1970
|
| [103] |
Brown BA, Springer B, Steingrube VA, et al. (1999) Mycobacterium wolinskyi sp. nov. and Mycobacterium goodii sp. nov., two new rapidly growing species related to Mycobacterium smegmatis and associated with human wound infections: a cooperative study from the International Working Group on Mycobacterial Taxonomy. Int J Syst Bacteriol 49 Pt 4: 1493-1511. doi: 10.1099/00207713-49-4-1493
|
| [104] |
Maya-Hoyos M, Leguizamon J, Marino-Ramirez L, et al. (2015) Sliding motility, biofilm formation, and glycopeptidolipid production in Mycobacterium colombiense Strains. Biomed Res Int 2015: 419549. doi: 10.1155/2015/419549
|
| [105] |
McMullen AR, Mattar C, Kirmani N, et al. (2015) Brown-pigmented Mycobacterium mageritense as a cause of prosthetic valve endocarditis and bloodstream infection. J Clin Microbiol 53: 2777-2780. doi: 10.1128/JCM.01041-15
|
| [106] | Hawkins JE, Falco EB (1976) Mycobacterium resembling Mycobacterium fortuitum that produces brown pigment. J Clin Microbiol 3: 453-455. |
| [107] |
Stormer RS, Falkinham JO (1989) Differences in antimicrobial susceptibility of pigmented and unpigmented colonial variants of Mycobacterium avium. J Clin Microbiol 27: 2459-2465. doi: 10.1128/JCM.27.11.2459-2465.1989
|
| [108] |
Falkinham JO (2009) Surrounded by mycobacteria: nontuberculous mycobacteria in the human environment. J Appl Microbiol 107: 356-367. doi: 10.1111/j.1365-2672.2009.04161.x
|
| [109] |
Falkinham JO (2013) Ecology of nontuberculous mycobacteria-where do human infections come from? Semin Respir Crit Care Med 34: 95-102. doi: 10.1055/s-0033-1333568
|
| [110] |
Lipner EM, Knox D, French J, et al. (2017) A geospatial epidemiologic analysis of nontuberculous Mycobacterial infection: an ecological study in Colorado. Ann Am Thorac Soc 14: 1523-1532. doi: 10.1513/AnnalsATS.201701-081OC
|
| [111] |
Bodmer T, Miltner E, Bermudez LE (2000) Mycobacterium avium resists exposure to the acidic conditions of the stomach. FEMS Microbiol Lett 182: 45-49. doi: 10.1111/j.1574-6968.2000.tb08871.x
|
| [112] |
Dawrs SN, Kautz M, Chan ED, et al. (2020) Mycobacterium abscessus and Gastroesophageal Reflux: An in vitro Study. Am J Respir Crit Care Med 202: 466-469. doi: 10.1164/rccm.202001-0011LE
|
| [113] |
Nikitushkin VD, Shleeva MO, Zinin AI, et al. (2016) The main pigment of the dormant Mycobacterium smegmatis is porphyrin. FEMS Microbiol Lett 363. doi: 10.1093/femsle/fnw206
|
| [114] |
Garvey M (2018) Mycobacterium avium subspecies paratuberculosis: A possible causative agent in human morbidity and risk to public health safety. Open Vet J 8: 172-181. doi: 10.4314/ovj.v8i2.10
|
| [115] | Nguyen Le T (2017) An investigation of the expression of pigment genes in Mycobacterium avium subspecies paratuberculosis, strain K10: University of Nottingham. |
| [116] |
Liu CI, Liu GY, Song Y, et al. (2008) A cholesterol biosynthesis inhibitor blocks Staphylococcus aureus virulence. Science 319: 1391-1394. doi: 10.1126/science.1153018
|
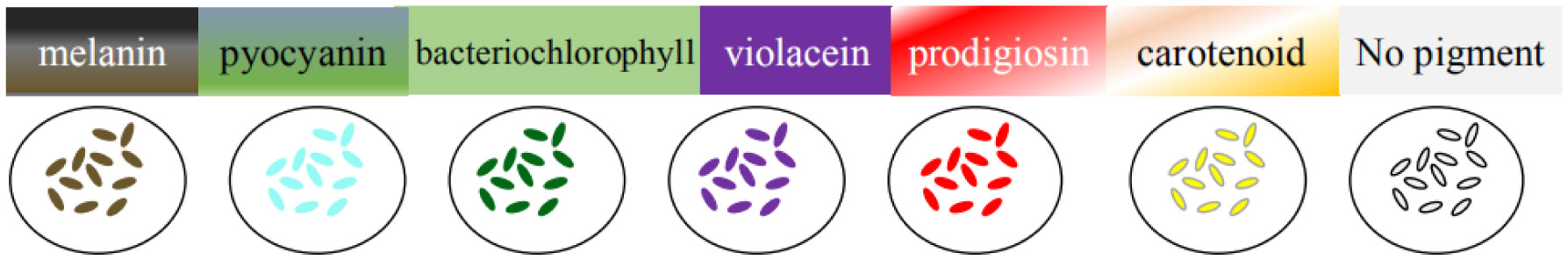
Figures(1) / Tables(2)

Tru Tran, Stephanie N. Dawrs, Grant J. Norton, Ravleen Virdi, Jennifer R. Honda. Brought to you courtesy of the red, white, and blue–pigments of nontuberculous mycobacteria[J]. AIMS Microbiology, 2020, 6(4): 434-450. doi: 10.3934/microbiol.2020026







 DownLoad:
DownLoad: