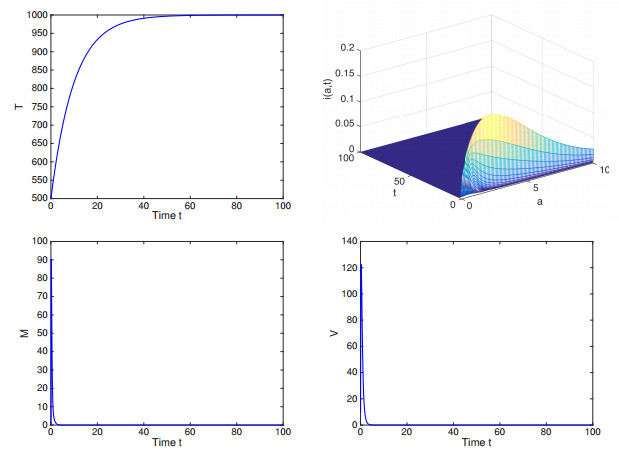
Recent studies reveal that pyroptosis is associated with the release of inflammatory cytokines which can attract more target cells to be infected. In this paper, a novel age-structured virus infection model incorporating cytokine-enhanced infection is investigated. The asymptotic smoothness of the semiflow is studied. With the help of characteristic equations and Lyapunov functionals, we have proved that both the local and global stabilities of the equilibria are completely determined by the threshold $ \mathcal{R}_0 $. The result shows that cytokine-enhanced viral infection also contributes to the basic reproduction number $ \mathcal{R}_0 $, implying that it may not be enough to eliminate the infection by decreasing the basic reproduction number of the model without considering the cytokine-enhanced viral infection mode. Numerical simulations are carried out to illustrate the theoretical results.
Citation: Jinhu Xu. Dynamic analysis of a cytokine-enhanced viral infection model with infection age[J]. Mathematical Biosciences and Engineering, 2023, 20(5): 8666-8684. doi: 10.3934/mbe.2023380
Recent studies reveal that pyroptosis is associated with the release of inflammatory cytokines which can attract more target cells to be infected. In this paper, a novel age-structured virus infection model incorporating cytokine-enhanced infection is investigated. The asymptotic smoothness of the semiflow is studied. With the help of characteristic equations and Lyapunov functionals, we have proved that both the local and global stabilities of the equilibria are completely determined by the threshold $ \mathcal{R}_0 $. The result shows that cytokine-enhanced viral infection also contributes to the basic reproduction number $ \mathcal{R}_0 $, implying that it may not be enough to eliminate the infection by decreasing the basic reproduction number of the model without considering the cytokine-enhanced viral infection mode. Numerical simulations are carried out to illustrate the theoretical results.

| [1] |
A. S. Perelson, A. Neumann, M. Markowitz, J. M. Leonard, D. D. Ho, HIV-1 dynamics in vivo: Virion clearance rate, infected cell life-span, and viral generation time, Science, 271 (1996), 158–286. https://doi.org/10.1126/science.271.5255.1582 doi: 10.1126/science.271.5255.1582

|
| [2] |
A. S. Perelson, P. W. Nelson, Mathematical analysis of HIV-1 dynamics in vivo, SIAM Rev., 41 (1999), 3–44. https://doi.org/10.1137/S0036144598335107 doi: 10.1137/S0036144598335107
|
| [3] |
J. Mittler, B. Sulzer, A. Neumann, A. S. Perelson, Influence of delayed virus production on viral dynamics in HIV-1 infected patients, Math. Biosci., 152 (1998), 143–163. https://doi.org/10.1016/S0025-5564(98)10027-5 doi: 10.1016/S0025-5564(98)10027-5
|
| [4] |
P. W. Nelson, J. Murray, A. S. Perelson, A model of HIV-1 pathogenesis that includes an intracellular delay, Math. Biosci., 163 (2000), 201–215. https://doi.org/10.1016/S0025-5564(99)00055-3 doi: 10.1016/S0025-5564(99)00055-3
|
| [5] |
P. de Leenheer, H. L. Smith, Virus dynamics: A global analysis, SIAM J. Appl. Math., 63 (2003), 1313–1327. https://doi.org/10.1137/S0036139902406905 doi: 10.1137/S0036139902406905
|
| [6] |
Y. Wang, Y. C. Zhou, Oscillatory viral dynamics in a delayed HIV pathogenesis model, Math. Biosic., 219 (2009), 104–112. https://doi.org/10.1016/j.mbs.2009.03.003 doi: 10.1016/j.mbs.2009.03.003
|
| [7] |
M. Y. Li, H. Shu, Global dynamics of an in-host viral model with intracellular delay, Bull. Math. Biol., 72 (2010), 1492–1505. https://doi.org/10.1007/s11538-010-9503-x doi: 10.1007/s11538-010-9503-x
|
| [8] |
D. S. Callaway, A. S. Perelson, HIV-1 infection and low steady state viral loads, Bull. Math. Biol., 64 (2002), 29–64. https://doi.org/10.1006/bulm.2001.0266 doi: 10.1006/bulm.2001.0266
|
| [9] |
Y. Shang, Analytical solution for an in-host viral infection model with time-inhomogeneous rates, Acta. Phys. Pol. B, 46 (2015), 1567–1577. https://doi.org/10.5506/APhysPolB.46.1567 doi: 10.5506/APhysPolB.46.1567
|
| [10] |
Y. Shang, Global stability of disease-free equilibria in a two-group SI model with feedback control, Nonlinear Anal. Model., 20 (2015), 501–508. https://doi.org/10.15388/NA.2015.4.3 doi: 10.15388/NA.2015.4.3
|
| [11] |
M. A. Gilchrist, D. Coombs, A. S. Perelson, Optimizing within-host viral fitness: Infected cell lifespan and virion production rate, J. Theor. Biol., 229 (2004), 281–288. https://doi.org/10.1016/j.jtbi.2004.04.015 doi: 10.1016/j.jtbi.2004.04.015
|
| [12] |
C. L. Althaus, A. S. de Vos, R. J. De Boer, Reassessing the human immunodeficiency virus type 1 life cycle through age-structured modeling: Life span of infected cells, viral generation time, and basic reproductive number, $r_0$, J. Virol., 83 (2009), 7659–7667. https://doi.org/10.1128/JVI.01799-08 doi: 10.1128/JVI.01799-08
|
| [13] |
P. W. Nelson, M. A. Gilchrist, D. Coombs, J. M. Hyman, A. Perelson, An age-structured model of HIV infection that allows for variations in the production rate of viral particles and the death rate of productively infected cells, Math. Biosci. Eng., 1 (2004), 267–288. https://doi.org/10.3934/mbe.2004.1.267 doi: 10.3934/mbe.2004.1.267
|
| [14] |
G. Huang, X. Liu, Y. Takeuchi, Lyapunov functions and global stability for age-structured HIV infection model, SIAM J. Appl. Math., 72 (2012), 25–38. https://doi.org/10.1137/110826588 doi: 10.1137/110826588
|
| [15] |
L. Rong, Z. Feng, A.S. Perelson, Mathematical analysis of age-structured HIV-1 dynamics with combination antiretroviral therapy, SIAM J. Appl. Math., 67 (2007), 731–756. https://doi.org/10.1137/060663945 doi: 10.1137/060663945
|
| [16] |
Y. Yang, S. Ruan, D. Xiao, Global stability of an age-structured virus dynamics model with Beddington-DeAngelis infection function, Math. Biosci. Eng., 12 (2015), 859–877. https://doi.org/10.3934/mbe.2015.12.859 doi: 10.3934/mbe.2015.12.859
|
| [17] |
S. Wang, X. Song, Global properties for an age-structured within-host model with Crowley-Martin functional response, Int. J. Biomath., 10 (2017), 1750030. https://doi.org/10.1142/S1793524517500309 doi: 10.1142/S1793524517500309
|
| [18] |
J. Wang, R. Zhang, T. Kuniya, Global dynamics for a class of age-infection HIV models with nonlinear infection rate, J. Math. Anal. Appl., 432 (2015), 289–313. https://doi.org/10.1016/j.jmaa.2015.06.040 doi: 10.1016/j.jmaa.2015.06.040
|
| [19] |
X. Wang, Y. Lou, X. Song, Age-structured within-host HIV dynamics with multiple target cells, Stud. Appl. Math., 138 (2017), 43–76. https://doi.org/10.1111/sapm.12135 doi: 10.1111/sapm.12135
|
| [20] | J. Wang, R. Zhang, T. Kuniya, Mathematical analysis for an age-structured HIV infection model with saturation infection rate, Electron. J. Differ. Equations, 2015 (2015), 1–19. |
| [21] |
L. Zou, S. Ruan, W. Zhang, An age-structured model for the transmission dynamics of hepatitis B, SIAM J. Appl. Math., 70 (2010), 3121–3139. https://doi.org/10.1137/090777645 doi: 10.1137/090777645
|
| [22] |
S. Liu, R. Zhang, On an age-structured hepatitis B virus infection model with HBV DNA-containing capsids, Bull. Malays. Math. Sci. Soc., 44 (2021), 1345–1370. https://doi.org/10.1007/s40840-020-01014-6 doi: 10.1007/s40840-020-01014-6
|
| [23] |
X. Tian, R. Xu, J. Lin, Mathematical analysis of an age-structured HIV-1 infection model with CTL immune response, Math. Biosci. Eng., 16 (2019), 7850–7882. https://doi.org/10.3934/mbe.2019395 doi: 10.3934/mbe.2019395
|
| [24] |
X. Duan, S. Yuan, K. Wang, Dynamics of a diffusive age-structured HBV model with saturating incidence, Math. Biosci. Eng., 13 (2016), 935–968. https://doi.org/10.3934/mbe.2016024 doi: 10.3934/mbe.2016024
|
| [25] |
J. Yang, X. Wang, Dynamics and asymptotical profiles of an age-structured viral infection model with spatial diffusion, Appl. Math. Comput., 360 (2019), 236–254. https://doi.org/10.1016/j.amc.2019.05.007 doi: 10.1016/j.amc.2019.05.007
|
| [26] |
J. Wang, R. Zhang, Y. Gao, Global threshold dynamics of an infection age-space structured HIV infection model with neumann boundary condition, J. Dyn. Differ. Equations, 2021 (2021), 1–33. https://doi.org/10.1007/s10884-021-10086-2 doi: 10.1007/s10884-021-10086-2
|
| [27] |
N. Martin, Q. Sattentau, Cell-to-cell HIV-1 spread and its implications for immune evasion, Curr. Opin. HIV AIDS., 4 (2009), 143–149. https://doi.org/10.1097/COH.0b013e328322f94a doi: 10.1097/COH.0b013e328322f94a
|
| [28] |
B. K. Titanji, M. Aasa-Chapman, D. Pillay, C. Jolly, Protease inhibitors effectively block cell-to-cell spread of HIV-1 between T cells, Retrovirology, 10 (2013), 161. https://doi.org/10.1186/1742-4690-10-161 doi: 10.1186/1742-4690-10-161
|
| [29] |
H. Sato, J. Orenstein, D. S. Dimitrov, M. Martin, Cell-to-cell spread of HIV-1 occurs with minutes and may not involve the participation of virus particles, Virology, 186 (1992), 712–724. https://doi.org/10.1016/0042-6822(92)90038-Q doi: 10.1016/0042-6822(92)90038-Q
|
| [30] |
D. S. Dimitrov, R. L. Willey, H. Sato, L. J. Chang, R. Blumenthal, M. A. Martin, Quantitation of human immunodeficiency virus type 1 infection kinetics, J. Virol., 67 (1993), 2182–2190. https://doi.org/10.1128/jvi.67.4.2182-2190.1993 doi: 10.1128/jvi.67.4.2182-2190.1993
|
| [31] |
M. Sourisseau, N. Sol-Foulon, F. Porrot, F. Blanchet, O. Schwartz, Inefficient human immunodeficiency virus replication in mobile lymphocytes, J. Virol., 81 (2007), 1000–1012. https://doi.org/10.1128/JVI.01629-06 doi: 10.1128/JVI.01629-06
|
| [32] |
X. L. Lai, X. F. Zou, Modeling HIV-1 virus dynamics with both virus-to-cell infection and cell-to-cell transmission, SIAM J. Appl. Math., 74 (2014), 898–917. https://doi.org/10.1137/130930145 doi: 10.1137/130930145
|
| [33] |
X. L. Lai, X. F. Zou, Modeling cell-to-cell spread of HIV-1 with logistic target cell growth, J. Math. Anal. Appl., 426 (2015), 563–584. https://doi.org/10.1016/j.jmaa.2014.10.086 doi: 10.1016/j.jmaa.2014.10.086
|
| [34] |
Y. Yang, L. Zou, S. Ruan, Global dynamics of a delayed within-host viral infection model with both virus-to-cell and cell-to-cell transmissions, Math. Biosci., 270 (2015), 183–191. https://doi.org/10.1016/j.mbs.2015.05.001 doi: 10.1016/j.mbs.2015.05.001
|
| [35] |
J. Wang, M. Guo, X. Liu, Z. Zhao, Threshold dynamics of HIV-1 virus model with cell-to-cell transmission, cell-mediated immune responses and distributed delay, Appl. Math. Comput., 291 (2016), 149–161. https://doi.org/10.1016/j.amc.2016.06.032 doi: 10.1016/j.amc.2016.06.032
|
| [36] |
J. Wang, J. Yang, T. Kuniya, Dynamics of a PDE viral infection model incorporating cell-to-cell tranmission, J. Math. Anal. Appl., 444 (2016), 1542–1564. https://doi.org/10.1016/j.jmaa.2016.07.027 doi: 10.1016/j.jmaa.2016.07.027
|
| [37] |
F. Li, J. Wang, Analysis of an HIV infection model with logistic target-cell growth and cell-to-cell transmission, Chaos Solitons Fractals, 81 (2015), 136–145. https://doi.org/10.1016/j.chaos.2015.09.003 doi: 10.1016/j.chaos.2015.09.003
|
| [38] | R. Zhang, S. Liu, Global dynamics of an age-structured within-host viral infection model with cell-to-cell transmission and general humoral immunity response, Math. Biosci. Eng., 17 2020, 1450–1478. https://doi.org/10.3934/mbe.2020075 |
| [39] |
W. Wang, X. Wang, K. Guo, W. Ma, Global analysis of a diffusive viral model with cell-to-cell infection and incubation period, Math. Method Appl. Sci., 43 (2020), 5963–5978. https://doi.org/10.1002/mma.6339 doi: 10.1002/mma.6339
|
| [40] |
X. Ren, Y. Tian, L. Liu, X. Liu, A reaction-diffusion within-host HIV model with cell-to-cell transmission, J. Math. Biol., 76 (2018), 1831–1872. https://doi.org/10.1007/s00285-017-1202-x doi: 10.1007/s00285-017-1202-x
|
| [41] |
Y. Xu, Z. Zhu, Y. Yang, F. Meng, Vectored immunoprophylaxis and cell-to-cell transmission in HIV dynamics, Int. J. Bifurcation Chaos, 30 (2020), 1–19. https://doi.org/10.1142/S0218127420501850 doi: 10.1142/S0218127420501850
|
| [42] |
T. Guo, Z. Qiu, L. Rong, Modeling the role of macrophages in HIV persistence during antiretroviral therapy, J. Math. Biol., 81 (2020), 369–402. https://doi.org/10.1007/s00285-020-01513-x doi: 10.1007/s00285-020-01513-x
|
| [43] |
Y. Gao, J. Wang, Threshold dynamics of a delayed nonlocal reaction-diffusion HIV infection model with both cell-free and cell-to-cell transmissions, J. Math. Anal. Appl., 488 (2020), 124047. https://doi.org/10.1016/j.jmaa.2020.124047 doi: 10.1016/j.jmaa.2020.124047
|
| [44] |
C. Y. Cheng, Y. Dong, Y. Takeuchi, An age-structured virus model with two routes of infection in heterogeneous environments, Nonlinear Anal. RWA., 39 (2018), 464–491. https://doi.org/10.1016/j.nonrwa.2017.07.013 doi: 10.1016/j.nonrwa.2017.07.013
|
| [45] |
J. Xu, Y. Geng, Y. Zhou, Global dynamics for an age-structured HIV virus infection model with cellular infection and antiretroviral therapy, Appl. Math. Comput., 305 (2017), 62–83. https://doi.org/10.1016/j.amc.2017.01.064 doi: 10.1016/j.amc.2017.01.064
|
| [46] |
G. Doitsh, N. Galloway, X. Geng, Z. Yang, K. M. Monroe, O. Zepeda, et al., Pyroptosis drives CD4 T-cell depletion in HIV-1 infection, Nature, 505 (2014), 509–514. https://doi.org/10.1038/nature12940 doi: 10.1038/nature12940
|
| [47] |
G. Doitsh, M. Cavrois, K.G. Lassen, O. Zepeda, Z. Yang, M. L. Santiago, et al., Abortive HIV infection mediates CD4 T cell depletion and inflammation in human lymphoid tissue, Cell, 143 (2010), 789–801. https://doi.org/10.1016/j.cell.2010.11.001 doi: 10.1016/j.cell.2010.11.001
|
| [48] |
A. L. Cox, R. F. Siliciano, HIV: Not-so-innocent bystanders, Nature, 505 (2014), 492–493. https://doi.org/10.1038/505492a doi: 10.1038/505492a
|
| [49] |
S. Wang, P. Hottz, M. Schechter, L. Rong, Modeling the slow CD4+ T cell decline in HIV-infected individuals, PLoS Comput. Biol., 11 (2015), 1–25. https://doi.org/10.1371/journal.pcbi.1004665 doi: 10.1371/journal.pcbi.1004665
|
| [50] |
W. Wang, T. Zhang, Caspase-1-mediated pyroptosis of the predominance for driving CD4+ T cells death: a nonlocal spatial mathematical model, Bull. Math. Biol., 80 (2018), 540–582. https://doi.org/10.1007/s11538-017-0389-8 doi: 10.1007/s11538-017-0389-8
|
| [51] | W. Wang, Z. Feng, Global dynamics of a diffusive viral infection model with spatial heterogeneity, Nonlinear Anal. RWA., 2022 2022, 103763. https://doi.org/10.1016/j.nonrwa.2022.103763 |
| [52] |
Y. Jiang, T. Zhang, Global stability of a cytokine-enhanced viral infection model with nonlinear incidence rate and time delays, Appl. Math. Letters., 132 (2022), 108110. https://doi.org/10.1016/j.aml.2022.108110 doi: 10.1016/j.aml.2022.108110
|
| [53] | P. Magal, S. Ruan, Theory And Applications of Abstract Semilinear Cauchy Problems, Springer International Publishing, 2018. |
| [54] | H. L. Smith, H. R. Thieme, Dynamical Systems and Population Persistence, American Mathematical Society, Providence, 2011. |
| [55] |
C. C. McCluskey, Global stability for an SEI epidemiological model with continuous age-structure in the exposed and infectious classes, Math. Biosci. Eng., 9 (2012), 819–841. https://doi.org/10.3934/mbe.2012.9.819 doi: 10.3934/mbe.2012.9.819
|
| [56] |
Y. Shang, The limit behavior of a stochastic logistic model with individual time-dependent rates, J. Math., 2013 (2013), 1–7. https://doi.org/10.1155/2013/502635 doi: 10.1155/2013/502635
|
| [57] | J. P. LaSalle, The Stability of Dynamical Systems, SIAM, 1976. ttps://doi.org/10.21236/ADA031020 |


Figures(3)

Jinhu Xu. Dynamic analysis of a cytokine-enhanced viral infection model with infection age[J]. Mathematical Biosciences and Engineering, 2023, 20(5): 8666-8684. doi: 10.3934/mbe.2023380







 DownLoad:
DownLoad: