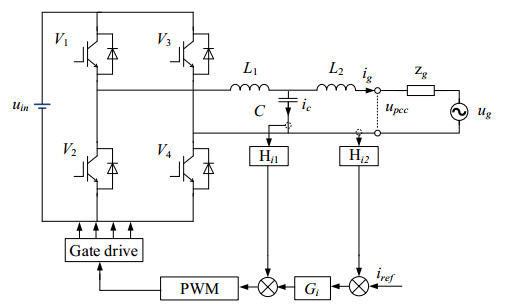
The regenerative braking in the tram allows the energy to be returned to the power grid through a power inverter. Since the inverter location between the tram and the power grid is not fixed, resulting in a wide variety of impedance networks at grid coupling points, posing a severe threat to the stable operation of the grid-tied inverter (GTI). By independently changing the loop characteristics of the GTI, the adaptive fuzzy PI controller (AFPIC) can adjust according to different impedance network parameters. It is challenging to fulfill the stability margin requirements of GTI under high network impedance since the PI controller has phase lag characteristics. A correction method of series virtual impedance is proposed, which connects the inductive link in a series configuration with the inverter output impedance, correcting the inverter equivalent output impedance from resistance-capacitance to resistance-inductance and improving the system stability margin. Feedforward control is adopted to improve the system's gain in the low-frequency band. Finally, the specific series impedance parameters are obtained by determining the maximum network impedance and setting the minimum phase margin of 45°. The realization of virtual impedance is simulated by conversion to an equivalent control block diagram, and the effectiveness and feasibility of the proposed method are verified by simulation and a 1 kW experimental prototype.
Citation: Sue Wang, Chaohong Zhou, Saleem Riaz, Xuanchen Guo, Haider Zaman, Alsharef Mohammad, Ahmad Aziz Al-Ahmadi, Yasser Mohammed Alharbi, NasimUllah. Adaptive fuzzy-based stability control and series impedance correction for the grid-tied inverter[J]. Mathematical Biosciences and Engineering, 2023, 20(2): 1599-1616. doi: 10.3934/mbe.2023073
The regenerative braking in the tram allows the energy to be returned to the power grid through a power inverter. Since the inverter location between the tram and the power grid is not fixed, resulting in a wide variety of impedance networks at grid coupling points, posing a severe threat to the stable operation of the grid-tied inverter (GTI). By independently changing the loop characteristics of the GTI, the adaptive fuzzy PI controller (AFPIC) can adjust according to different impedance network parameters. It is challenging to fulfill the stability margin requirements of GTI under high network impedance since the PI controller has phase lag characteristics. A correction method of series virtual impedance is proposed, which connects the inductive link in a series configuration with the inverter output impedance, correcting the inverter equivalent output impedance from resistance-capacitance to resistance-inductance and improving the system stability margin. Feedforward control is adopted to improve the system's gain in the low-frequency band. Finally, the specific series impedance parameters are obtained by determining the maximum network impedance and setting the minimum phase margin of 45°. The realization of virtual impedance is simulated by conversion to an equivalent control block diagram, and the effectiveness and feasibility of the proposed method are verified by simulation and a 1 kW experimental prototype.

| [1] |
Y. He, K. M. Kockelman, K. A. Perrine, Optimal locations of US fast charging stations for long-distance trip completion by battery electric vehicles, J. Cleaner Prod., 214 (2019), 452–461. https://doi.org/10.1016/j.jclepro.2018.12.188 doi: 10.1016/j.jclepro.2018.12.188

|
| [2] |
V. A. Kleftakis, N. D. Hatziargyriou, Optimal control of reversible substations and wayside storage devices for voltage stabilization and energy savings in metro railway networks, IEEE Trans. Transp. Electrif., 5 (2019), 515–523. https://doi.org/10.1109/TTE.2019.2913355 doi: 10.1109/TTE.2019.2913355
|
| [3] |
E. Samavati, H. R. Mohammadi, An improved method for harmonic mitigation and stability improvement of the grid-connected inverters under local load variation and weak grid condition, Int J. Electr. Power Energy Syst., 123 (2020), 106310. https://doi.org/10.1016/j.ijepes.2020.106310 doi: 10.1016/j.ijepes.2020.106310
|
| [4] | S. Fu, X. Zhang, D. Xu, Improved control method of grid-connected converter based on voltage perturbation compensation under weak grid conditions, in 2019 10th International Conference on Power Electronics and ECCE Asia (ICPE 2019-ECCE Asia), (2019), 2586–2591. https://doi.org/10.23919/ICPE2019-ECCEAsia42246.2019.8797350 |
| [5] |
F. Hao, G. Zhang, J. Chen, Z. Liu, D. Xu, Y. Wang, et al., Optimal voltage regulation and power sharing in traction power systems with reversible converters, IEEE Trans. Power Syst., 35 (2020), 2726–2735. https://doi.org/10.1109/TPWRS.2020.2968108 doi: 10.1109/TPWRS.2020.2968108
|
| [6] |
Y. Wang, W. Wen, C. Wang, H. Liu, X. Zhan, X. Xiao, et al., Adaptive voltage droop method of multiterminal VSC-HVDC systems for DC voltage deviation and power sharing, IEEE Trans. Power Del., 34 (2020), 169–176. https://doi.org/10.1109/TPWRD.2018.2844330 doi: 10.1109/TPWRD.2018.2844330
|
| [7] |
B. Liu, H. Ben, Xi. Zhang, T. Meng, X. Wang, Stabilization of a cascaded AC–DC–DC system with small bus capacitance based on small signal analysis, IEEE Trans. Ind. Appl., 54 (2018), 4650–4659. https://doi.org/10.1109/TIA.2018.2837042 doi: 10.1109/TIA.2018.2837042
|
| [8] |
W. Zhang, Y. Fang, R. Ye, Z. Wang, Analysis and design of a double fuzzy PI controller of a voltage outer loop in a reversible three-phase PWM converter, Energies, 13 (2020), 3778. https://doi.org/10.3390/en13153778 doi: 10.3390/en13153778
|
| [9] |
T. Hou, C. Y. Zhang, H. X. Niu, Quasi-Z source inverter control of PV grid-connected based on fuzzy PCI, J. Elect. Sci. Technol., 19 (2021), 100021. https://doi.org/10.1016/j.jnlest.2020.100021 doi: 10.1016/j.jnlest.2020.100021
|
| [10] |
C. W. Tao, C. M. Wang, C. W. Chang, A design of a DC-AC inverter using a modified ZVS-PWM auxiliary commutation pole and a DSP-based PID-like fuzzy control, IEEE Trans. Ind. Electron., 63 (2015), 397–405. https://doi.org/10.1109/TIE.2015.2477061 doi: 10.1109/TIE.2015.2477061
|
| [11] |
Y. Dai, L. Zhang, G. Liu, C. Yang, D. Zhang, X. Huang, Prescribed-performance based finite-time adaptive fuzzy control for PV inverter in islanded systems, Int. J. Electron. Power Energy Syst., 133 (2021), 107254. https://doi.org/10.1016/j.ijepes.2021.107254 doi: 10.1016/j.ijepes.2021.107254
|
| [12] |
Z. Li, W. Cheng, Q. Xiao, W. Liao, Deadbeat predictive power control with fuzzy PI compound controller and power predictive corrector for PWM rectifier under unbalanced grid conditions, Int. J. Fuzzy Syst., 22 (2020), 1277–1288. https://doi.org/10.1007/s40815-020-00847-4 doi: 10.1007/s40815-020-00847-4
|
| [13] |
T. Lei, S. Riaz, N. Zanib, M. Batool, F. Pan, S. Zhang, Performance analysis of grid-connected distributed generation system integrating a hybrid wind-pv farm using UPQC, Complexity, 2022 (2022), 14. https://doi.org/10.1155/2022/4572145 doi: 10.1155/2022/4572145
|
| [14] |
Z. Zheng, N. Wang, Z. Sun, Fuzzy PI compound control of PWM rectifiers with applications to marine vehicle electric propulsion system, Int. J. Fuzzy Syst., 20 (2018), 587–596. https://doi.org/10.1007/s40815-017-0394-y doi: 10.1007/s40815-017-0394-y
|
| [15] |
S. Riaz, L. Hui, M. S. Aldemir, F. Afzal, A future concern of iterative learning control: A survey. journal of statistics and management systems, 24 (2021), 1301–1322. https://doi.org/10.1080/09720510.2020.1856421 doi: 10.1080/09720510.2020.1856421
|
| [16] |
R. Wang, Q. Sun, W. Hu, J. Xiao, H. Zhang, P. Wang, Stability-oriented droop coefficients region identification for inverters within weak grid: an impedance-based approach, IEEE Trans. Syst., 51 (2020), 2258–2268. https://doi.org/10.1109/TSMC.2020.3034243 doi: 10.1109/TSMC.2020.3034243
|
| [17] |
Z. Liu, J. Liu, W. Bao, Y. Zhao, Infinity-norm of impedance-based stability criterion for three-phase AC distributed power systems with constant power loads, IEEE Trans. Power Electron., 30 (2014), 3030–3043. https://doi.org/10.1109/TPEL.2014.2331419 doi: 10.1109/TPEL.2014.2331419
|
| [18] |
J. Xu, S. Xie, LCL-resonance damping strategies for grid-connected inverters with LCL filters: A comprehensive review, J. Mod. Power Syst. Clean Energy, 6 (2018), 292–305. https://doi.org/10.1007/s40565-017-0319-7 doi: 10.1007/s40565-017-0319-7
|
| [19] |
S. Li, X. Li, X. Lee, Weighted average current method for active damping control based on grid voltage feed-forward, J. Cloud Comput., 10 (2021), 1–12. https://doi.org/10.1186/s13677-021-00236-8 doi: 10.1186/s13677-021-00236-8
|
| [20] |
L. Li, W. Chen, Y. Han, Q. Li, Y. Pu, A stability enhancement method based on adaptive virtual resistor for electric-hydrogen hybrid DC microgrid grid-connected inverter under weak grid, Electr. Power Syst. Res., 191 (2021), 106882. https://doi.org/10.1016/j.epsr.2020.106882 doi: 10.1016/j.epsr.2020.106882
|
| [21] |
Z. Lin, X. Ruan, L. Wu, H. Zhang, W. Li, Multi resonant component-based grid-voltage-weighted feedforward scheme for grid-connected inverter to suppress the injected grid current harmonics under weak grid, IEEE Trans. Power Electron., 35 (2020), 9784–9793. https://doi.org/10.1109/TPEL.2020.2970514 doi: 10.1109/TPEL.2020.2970514
|
| [22] | Z. Xu, A. Xu, S. Xie, Dual-loop grid current control technique for grid-connected inverter using an LCL filter, Proc. Chin. Soc. Elect. Eng., 29 (2009), 36–41. |
| [23] | Y. N. Wang, C. Y. Guo, S. Yang, Non-monotonic characteristics of stability margin of MMC system with the change of inner loop current control bandwidth and its mechanism analysis, Proc. Chin. Soc. Elect. Eng., 42 (2022), 3538–3548. |
| [24] |
M. F. M. Arani, Y. A. Mohamed, Analysis and performance enhancement of vector-controlled VSC in HVDC links connected to very weak grids, IEEE Trans. Power Syst., 32 (2017), 684–693. https://doi.org/10.1109/TPWRS.2016.2540959 doi: 10.1109/TPWRS.2016.2540959
|
| [25] |
D. H. Sun, S. Y. Xu, T. Xu, Q. Guo, J. He, B. Zhao, et al., Definition and index of short circuit ratio for multiple renewable energy stations, Proc. Chin. Soc. Elect. Eng., 41 (2021), 497–506. https://doi.org/10.13334/j.0258-8013.pcsee.202112 doi: 10.13334/j.0258-8013.pcsee.202112
|
| [26] |
T. Xue, P. Sun, Z. Xu, Q. Luo, Feedforward phase compensation method of LCL grid-connected inverter based on all-pass filter in weak grid, IET Power Electron., 13 (2020), 4407–4416. https://doi.org/10.1049/iet-pel.2020.0897 doi: 10.1049/iet-pel.2020.0897
|
| [27] |
D. Yang, X. Ruan, H. Wu, A virtual impedance method to improve the performance of LCL-type grid-connected inverters under weak grid conditions, Proc. Chin. Soc. Elect. Eng., 34 (2014), 2327–2335. https://doi.org/10.13334/j.0258-8013.pcsee.2014.15.001 doi: 10.13334/j.0258-8013.pcsee.2014.15.001
|


















Figures(19) / Tables(2)

Sue Wang, Chaohong Zhou, Saleem Riaz, Xuanchen Guo, Haider Zaman, Alsharef Mohammad, Ahmad Aziz Al-Ahmadi, Yasser Mohammed Alharbi, NasimUllah. Adaptive fuzzy-based stability control and series impedance correction for the grid-tied inverter[J]. Mathematical Biosciences and Engineering, 2023, 20(2): 1599-1616. doi: 10.3934/mbe.2023073







 DownLoad:
DownLoad: