Within the framework of physical and ecological integrated control of cyanobacteria bloom, because the outbreak of cyanobacteria bloom can form cyanobacteria clustering phenomenon, so a new aquatic ecological model with clustering behavior is proposed to describe the dynamic relationship between cyanobacteria and potential grazers. The biggest advantage of the model is that it depicts physical spraying treatment technology into the existence pattern of cyanobacteria, then integrates the physical and ecological integrated control with the aggregation of cyanobacteria. Mathematical theory works mainly investigate some key threshold conditions to induce Transcritical bifurcation and Hopf bifurcation of the model $ (2.1) $, which can force cyanobacteria and potential grazers to form steady-state coexistence mode and periodic oscillation coexistence mode respectively. Numerical simulation works not only explore the influence of clustering on the dynamic relationship between cyanobacteria and potential grazers, but also dynamically show the evolution process of Transcritical bifurcation and Hopf bifurcation, which can be clearly seen that the density of cyanobacteria decreases gradually with the evolution of bifurcation dynamics. Furthermore, it should be worth explaining that the most important role of physical spraying treatment technology can break up clumps of cyanobacteria in the process of controlling cyanobacteria bloom, but cannot change the dynamic essential characteristics of cyanobacteria and potential grazers represented by the model $ (2.1) $, this result implies that the physical spraying treatment technology cannot fundamentally eliminate cyanobacteria bloom. In a word, it is hoped that the results of this paper can provide some theoretical support for the physical and ecological integrated control of cyanobacteria bloom.
Citation: Qiulin Huang, Hengguo Yu, Chuanjun Dai, Zengling Ma, Qi Wang, Min Zhao. Dynamic analysis of a new aquatic ecological model based on physical and ecological integrated control[J]. Mathematical Biosciences and Engineering, 2023, 20(1): 930-954. doi: 10.3934/mbe.2023043
Within the framework of physical and ecological integrated control of cyanobacteria bloom, because the outbreak of cyanobacteria bloom can form cyanobacteria clustering phenomenon, so a new aquatic ecological model with clustering behavior is proposed to describe the dynamic relationship between cyanobacteria and potential grazers. The biggest advantage of the model is that it depicts physical spraying treatment technology into the existence pattern of cyanobacteria, then integrates the physical and ecological integrated control with the aggregation of cyanobacteria. Mathematical theory works mainly investigate some key threshold conditions to induce Transcritical bifurcation and Hopf bifurcation of the model $ (2.1) $, which can force cyanobacteria and potential grazers to form steady-state coexistence mode and periodic oscillation coexistence mode respectively. Numerical simulation works not only explore the influence of clustering on the dynamic relationship between cyanobacteria and potential grazers, but also dynamically show the evolution process of Transcritical bifurcation and Hopf bifurcation, which can be clearly seen that the density of cyanobacteria decreases gradually with the evolution of bifurcation dynamics. Furthermore, it should be worth explaining that the most important role of physical spraying treatment technology can break up clumps of cyanobacteria in the process of controlling cyanobacteria bloom, but cannot change the dynamic essential characteristics of cyanobacteria and potential grazers represented by the model $ (2.1) $, this result implies that the physical spraying treatment technology cannot fundamentally eliminate cyanobacteria bloom. In a word, it is hoped that the results of this paper can provide some theoretical support for the physical and ecological integrated control of cyanobacteria bloom.

| [1] |
H. Xu, H. Jiang, G. Yu, L. Yang, Towards understanding the role of extracellular polymeric substances in cyanobacterial Microcystis aggregation and mucilaginous bloom formation, Chemosphere, 117 (2014), 815–822. https://doi.org/10.1016/j.chemosphere.2014.10.061 doi: 10.1016/j.chemosphere.2014.10.061

|
| [2] |
B. Q. Qin, G. J. Yang, J. R. Ma, J. M. Deng, W. Li, T. Wu, et al., Dynamics of variability and mechanism of harmful cyanobacteria bloom in Lake Taihu, China, Chin. Sci. Bull., 61 (2016), 759–770. http://dx.doi.org/10.1360/N972015-00400 doi: 10.1360/N972015-00400
|
| [3] |
H. Z. Chen, S. P. Zuo, B. Q. Qin, L. T. Ye, H. Wang, Research progress in mechanism of Microcystis aggregation and migration, Environ. Sci. Technol. China, 42 (2019), 142–149. http://dx.doi.org/10.19672/j.cnki.1003-6504.2019.01.021 doi: 10.19672/j.cnki.1003-6504.2019.01.021
|
| [4] | F. X. Kong, R. H. Ma, J. F. Gao, X. Wang, The theory and practice of prevention, forecast and warning on cyanobacteria bloom in Lake Taihu, J. Lake Sci., 21 (2009), 314–328. |
| [5] |
F. A. Kibuye, A. Zzmyadi, E. C. Wert, A critical review on operation and performance of source water control strategies for cyanobacterial blooms: Part I-chemical control methods, Harmful Algae, 109 (2021), 102099. https://doi.org/10.1016/j.hal.2021.102099 doi: 10.1016/j.hal.2021.102099
|
| [6] |
F. A. Kibuye, A. Zzmyadi, E. C. Wert, A critical review on operation and performance of source water control strategies for cyanobacterial blooms: Part II-mechanical and biological control methods, Harmful Algae, 109 (2021), 102119. https://doi.org/10.1016/j.hal.2021.102119 doi: 10.1016/j.hal.2021.102119
|
| [7] |
L. Y. Yang, X. Y. Yang, L. M. Ren, X. Qian, L. Xiao, Mechanism and control strategy of cyanobacterial bloom in Lake Taihu, J. Lake Sci., 31 (2019), 18–27. http://dx.doi.org/10.18307/2019.0102 doi: 10.18307/2019.0102
|
| [8] |
T. Gorham, Y. Jia, C. K. Shum, J. Lee, Ten-year survey of cyanobacterial blooms in Ohio's waterbodies using satellite remote sensing, Harmful Algae, 66 (2017), 13–19. http://dx.doi.org/10.1016/j.hal.2017.04.013 doi: 10.1016/j.hal.2017.04.013
|
| [9] |
H. W. Paerl, T. G. Otten, Harmful cyanobacterial blooms: causes, consequences, and controls, Microb. Ecol., 65 (2013), 995–1010. http://dx.doi.org/10.1007/s00248-012-0159-y doi: 10.1007/s00248-012-0159-y
|
| [10] |
H. W. Paerl, Nuisance phytoplankton blooms in coastal, estuarine, and inland waters, Limnol Oceanogr, 33 (1988), 823–847. http://dx.doi.org/10.4319/lo.1988.33.4-part-2.0823 doi: 10.4319/lo.1988.33.4-part-2.0823
|
| [11] |
Z. Yang, F. X. Kong, X. L. Shi, H. Cao, Morphological response of Microcystis aeruginosa to grazing by different sorts of zooplankton, Hydrobiologia, 563 (2006), 225–230. http://dx.doi.org/10.1007/s1075–005-0008-9 doi: 10.1007/s1075–005-0008-9
|
| [12] |
U. Burkert, P. Hyenstrand, S. Drakare, P. Blomqvist, Effects of the mixotrophic flagellate Ochromonas sp. on colony formation in Microcystis aeruginosa, Aquat. Ecol., 35 (2001), 11–17. http://dx.doi.org/10.1023/a:1011454313607 doi: 10.1023/a:1011454313607
|
| [13] |
P. Chow-Fraser, D. O. Trew, D. Findlay, M. Stainton, A test of hypotheses to explain the sigmoidal relationship between total phosphorus and chlorophyll a concentrations in Canadian lakes, Can. J. Fish. Aquat. Sci., 51 (1994), 2052–2065. http://dx.doi.org/10.1139/f94-208 doi: 10.1139/f94-208
|
| [14] |
H. L. Gao, X. Qian, H. F. Wu, H. Li, H. Pan, C. Han, et al., Combined effects of submerged macrophytes and aquatic animals on the restoration of a eutrophic water body-a case study of Gonghu Bay, Harmful Algae, 102 (2017), 15–23. https://doi.org/10.1016/j doi: 10.1016/j
|
| [15] |
P. I. Boon, S. E. Bunn, J. D. Green, R. J. Shiel, Consumption of cyanobacteria by freshwater zooplankton: Implications for the success of 'top-down' control of cyanobacterial blooms in australia, Mar. Freshwater Res., 45 (1994), 875–887. https://doi.org/10.1071/MF9940875 doi: 10.1071/MF9940875
|
| [16] |
I. Setlíkova, S. Maciarzova, M. Blaha, T. Policar, Silver carp (Hypophthalmichthys molitrix) can non-mechanically digest cyanobacteria, Fish Physiol. Biochem., 46 (2020), 771–776. https://doi.org/10.1007/s10695-019-00751-1 doi: 10.1007/s10695-019-00751-1
|
| [17] |
H. Joshi, B. K. Jha, Chaos of calcium diffusion in Parkinson's infectious disease model and treatment mechanism via Hilfer fractional derivative, Math. Modell. Numer. Simul. Appl., 1 (2021), 84–94. https://doi.org/10.53391/mmnsa.2021.01.008 doi: 10.53391/mmnsa.2021.01.008
|
| [18] | H. Joshi, B. K. Jha, Modeling the spatiotemporal intracellular calcium dynamics in nerve cell with strong memory effects, Int. J. Nonlinear Sci. Numer. Simul., 2021 (2021). https://doi.org/10.1515/ijnsns-2020-0254 |
| [19] |
H. Joshi, B. K. Jha, Generalized diffusion characteristics of calcium model with concentration and memory of cells: A spatiotemporal approach, Iran. J. Sci. Technol. Trans. A Sci., 46 (2022), 309–322. https://doi.org/10.1007/s40995-021-01247-5 doi: 10.1007/s40995-021-01247-5
|
| [20] |
P. Veeresha, M. Yavuz, C. Baishya, A computational approach for shallow water forced Korteweg–De Vries equation on critical flow over a hole with three fractional operators, Int. J. Optim. Control Theor. Appl., 11 (2021), 52–67. https://doi.org/10.11121/ijocta.2021.1177 doi: 10.11121/ijocta.2021.1177
|
| [21] |
P. A. Naik, K. M. Owolabi, M. Yavuz, J. Zu, Chaotic dynamics of a fractional order HIV-1 model involving AIDS-related cancer cells, Chaos Solitons Fractals, 140 (2020), 110272. https://doi.org/10.1016/j.chaos.2020.110272 doi: 10.1016/j.chaos.2020.110272
|
| [22] |
M. Yavuz, N. Ozdemir, Analysis of an epidemic spreading model with exponential decay law, Math. Sci. Appl. E Notes, 8 (2020), 142–154. https://doi.org/10.36753/mathenot.691638 doi: 10.36753/mathenot.691638
|
| [23] |
M. Id, R. Khoshsiar, Z. Eskandari, Three-dimensional fractional system with the stability condition and chaos control, Math. Modell. Numer. Simul. Appl., 2 (2022), 41–47. https://doi.org/10.53391/mmnsa.2022.01.004 doi: 10.53391/mmnsa.2022.01.004
|
| [24] |
D. Anwarud, M. Abidin, Analysis of fractional-order vaccinated Hepatitis-B epidemic model with Mittag-Leffler kernels, Math. Modelling Numer. Simul. Appl., 2 (2022), 59–72. https://doi.org/10.53391/mmnsa.2022.006 doi: 10.53391/mmnsa.2022.006
|
| [25] |
P. A. Naik, Z. Eskandari, Z. Avazzadeh, J, Zu, Multiple bifurcations of a discrete-time prey-predator model with mixed functional response, Int. J. Bifurcation Chaos, 32 (2022), 2250050. https://doi.org/10.1142/S021812742250050X doi: 10.1142/S021812742250050X
|
| [26] |
P.A. Naik, Z. Eskandari, H. E. Shahkari, Flip and generalized flip bifurcations of a two-dimensional discrete-time chemical model, Math. Modell. Numer. Simul. Appl., 1 (2021), 95–101. https://doi.org/10.53391/mmnsa.2021.01.009 doi: 10.53391/mmnsa.2021.01.009
|
| [27] |
P. A. Naik, Z. Eskandari, M. Yavuz, J. Zu, Complex dynamics of a discrete-time Bazykin-Berezovskaya prey-predator model with a strong Allee effect, J. Comput. Appl. Math., 413 (2022), 114401. https://doi.org/10.1016/j.cam.2022.114401 doi: 10.1016/j.cam.2022.114401
|
| [28] | L. V. Bertalanffy, Theoretische Biologie, Springer, Berlin, 1932. https://doi.org/10.1007/978-3-662-36634-9 |
| [29] | S. E. Jorgensen, G. Bendoricchio, Fundamentals of Ecological Modelling, 3$^{nd}$ edition, Elsevier, 2001. |
| [30] |
S. N. Chen, X. L. Chen, Y. Peng, K, Peng, A mathematical model of the effect of nitrogen and phosphorus on the growth of blue-green algae population, Appl. Math. Modell., 33 (2009), 1097–1106. https://doi.org/10.1016/j.apm.2008.01.001 doi: 10.1016/j.apm.2008.01.001
|
| [31] |
Z. Rupawalla, N. Robinson, S. Schmidt, S. Li, S. Carruthers, E. Buisset, et al., Algae biofertilisers promote sustainable food production and a circular nutrient economy–An integrated empirical-modelling study, Sci. Total Environ., 796 (2021), 148913. https://doi.org/10.1016/j.scitotenv.2021.148913 doi: 10.1016/j.scitotenv.2021.148913
|
| [32] |
M. Sagehashi, A. Sakoda, M. Suzuki, A mathematical model of a shallow and Eutrophic Lake (The Keszthely Basin, Lake Balaton) and simulation of restorative manipulations, Water Res., 35 (2001), 1675–1686. https://doi.org/10.1016/S0043-1354(00)00435-8 doi: 10.1016/S0043-1354(00)00435-8
|
| [33] |
H. Y. Liu, H. G. Yu, C. J. Dai, Z. Ma, Q. Wang, M. Zhao, Dynamical analysis of an aquatic amensalism model with non-selective harvesting and Allee effect, Math. Biosci. Eng., 18 (2021), 8857–8882. https://doi.org/10.3934/mbe.2021437 doi: 10.3934/mbe.2021437
|
| [34] |
M. Sagehashi, A. Sakoda, M. Suzuki, A predictive model of long-term stability after biomanipulation of shallow lakes, Water Res., 34 (2000), 4014–4028. https://doi.org/10.1016/S0043-1354(00)00165-2 doi: 10.1016/S0043-1354(00)00165-2
|
| [35] |
X. X. Li, H. G. Yu, C.J. Dai, Z. Ma, Q. Wang, M. Zhao, Bifurcation analysis of a new aquatic ecological model with aggregation effect, Math. Comput. Simul., 190 (2021), 75–96. https://doi.org/10.1016/j.matcom.2021.05.015 doi: 10.1016/j.matcom.2021.05.015
|
| [36] |
A. Fadel, N. Sharaf, M. Siblini, K. Slim, A. Kobaissi, A simple modelling approach to simulate the effect of different climate scenarios on toxic cyanobacterial bloom in a eutrophic reservoir, Ecohydrol. Hydrobiol., 19 (2019), 359–369. https://doi.org/10.1016/j.ecohyd.2019.02.005 doi: 10.1016/j.ecohyd.2019.02.005
|
| [37] |
H. Serizawa, T. Amemiya, K. Itoh, Effects of buoyancy, transparency and zooplankton feeding on surface maxima and deep maxima: Comprehensive mathematical model for vertical distribution in cyanobacterial biomass, Ecol. Modell., 221 (2010), 2028–2037. https://doi.org/10.1016/j.ecolmodel.2010.05.009 doi: 10.1016/j.ecolmodel.2010.05.009
|
| [38] |
H. Serizawa, T. Amemiya, K. Itoh, Patchiness and bistability in the comprehensive cyanobacterial model (CCM), Ecol. Modell., 220 (2009), 764–773. https://doi.org/10.1016/j.ecolmodel.2008.12.015 doi: 10.1016/j.ecolmodel.2008.12.015
|
| [39] |
N. H. Gazi, K. Das, Structural stability analysis of an algal bloom mathematical model in tropic interaction, Nonlinear Anal. Real World Appl., 11 (2010), 2191–2206. https://doi.org/10.1016/j.nonrwa.2009.06.009 doi: 10.1016/j.nonrwa.2009.06.009
|
| [40] |
Y. S. Zhang, H. Y. Li, F. X. Kong, Y. Yu, M. Zhang, Role of colony intercelar space in the cyanobacteria bloom-forming, Environ. Sci., 32 (2011), 1602–1607. https://doi.org/10.13227/j.hjkx.2011.06.002 doi: 10.13227/j.hjkx.2011.06.002
|
| [41] |
S. Pereira, A. Zille, E. Micheletti, P. Moradas-Ferreira, R. De Philippis, P. Tamagnini, Complexity of cyanobacterial exopoly saccharides: composition, structures, inducing factors and putative genes involved in their biosynthesis and assembly, FEMS Microbiol. Rev., 33 (2009), 917–941. https://doi.org/10.1111/j.1574-6976.2009.00183.x doi: 10.1111/j.1574-6976.2009.00183.x
|
| [42] |
S. Xu, Y. J. Yang, J.Z. Xu, J. Shi, L. Song, Z. Wu, Physiological responses of populations and unicellular microcystis to shortterm high light stress, Acta Hydrobiol. Sinica, 41 (2017), 443–447. https://doi.org/10.7541/2017.55 doi: 10.7541/2017.55
|
| [43] |
Z. Yang, F. X. Kang, X. L. Shi, H. Cao, Morphological response of Microcystis aeruginosa to grazing by different sorts of zooplankton, Hydrobiologia, 563 (2006), 225–230. https://doi.org/10.1007/s10750-005-0008-9 doi: 10.1007/s10750-005-0008-9
|
| [44] |
H. Shen, L. R. Song, Comparative studies on physiological responses to phosphorus in two phenotype of bloom-forming microcystis, Hydrobiologia, 592 (2007), 475–786. https://doi.org/10.1007/s10750-007-0794-3 doi: 10.1007/s10750-007-0794-3
|
| [45] |
X. H. Wu, Z. X. Wu, L. R. Song, Phenotype and temperature affect the affinity for dissolved inorganic carbon in a cyanobacterium Microcystis, Hydrobiologia, 675 (2011), 175-186. https://doi.org/10.1007/s10750-011-0815-0 doi: 10.1007/s10750-011-0815-0
|
| [46] |
P. Xie, J. K. Liu, Practical success of biomanipulation using filter-feeding fish to control cyanobacteria blooms: A synthesis of decades of research and application in a subtropical hypereutrophic lake, Sci. World J., 1 (2014), 337–356. https://doi.org/10.1100/tsw.2001.67 doi: 10.1100/tsw.2001.67
|
| [47] |
H. H. Jakobsen, K. W. Tang, Effects of protozoan grazing on colony formation in Phaeocystis globosa (Prymnesiophyceae) and the potential costs and benefits, Aquat. Microb. Ecol., 27 (2002), 261–273. https://doi.org/10.3354/ame027261 doi: 10.3354/ame027261
|
| [48] |
K. W. Tang, Grazing and colony size development in Phaeocystis globosa (Prymnesiophyceae): the role of chemical signal, J. Plankton Res., 25 (2003), 831–842. https://doi.org/10.1093/plankt/25.7.831 doi: 10.1093/plankt/25.7.831
|
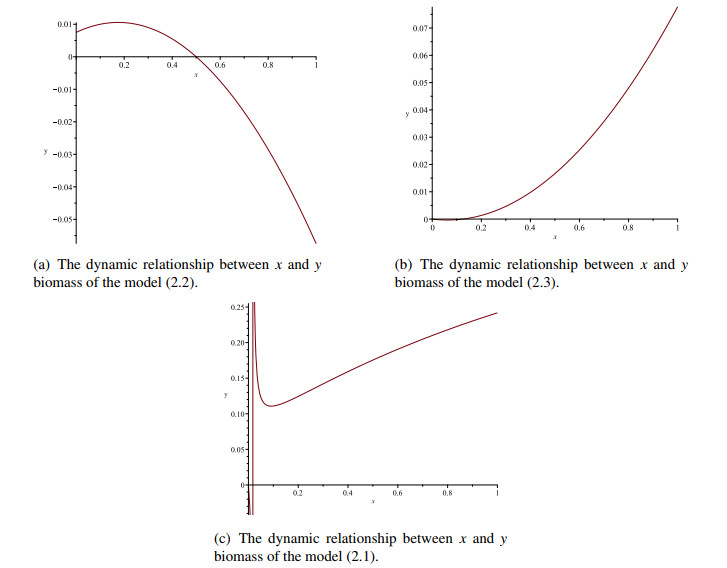
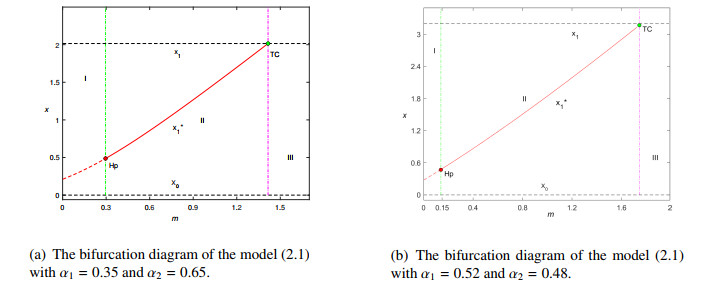
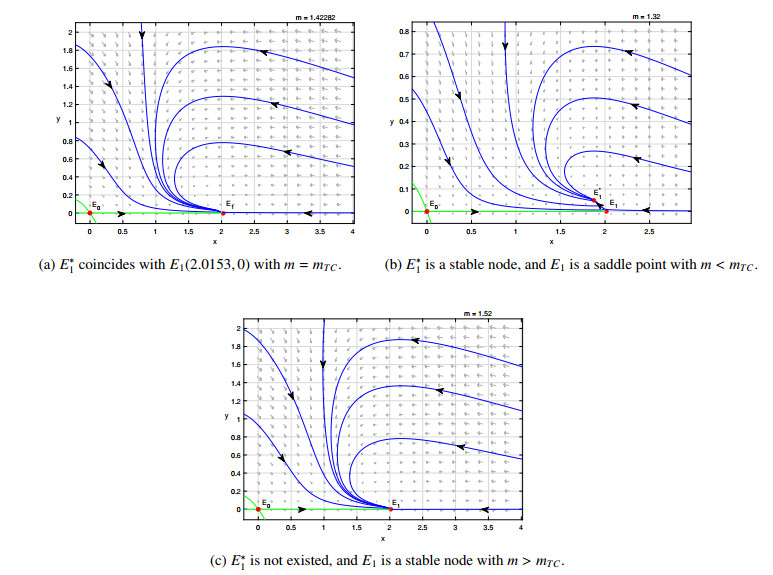
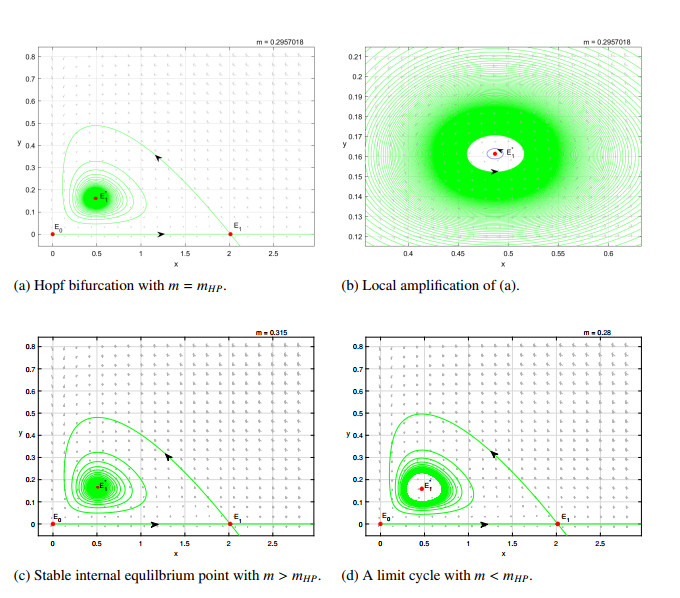
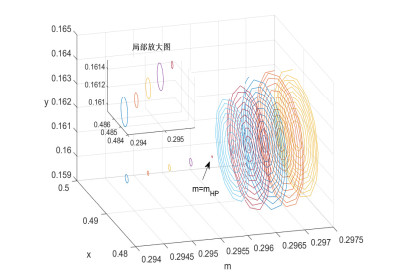
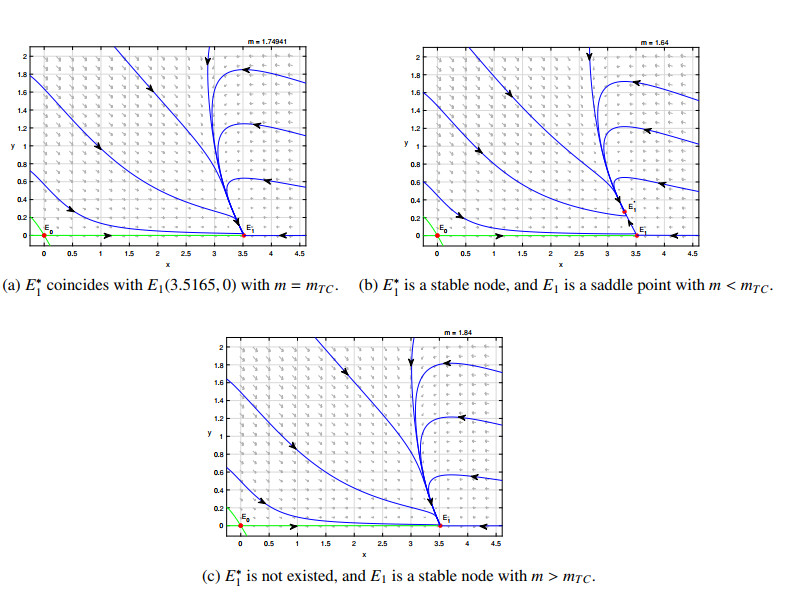
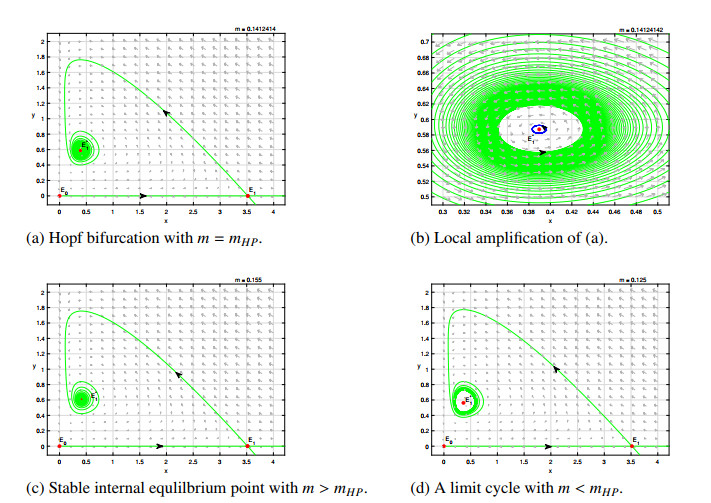
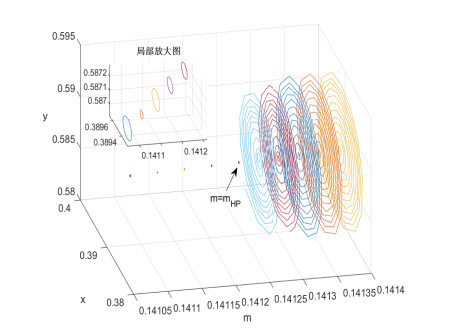
Figures(8)

Qiulin Huang, Hengguo Yu, Chuanjun Dai, Zengling Ma, Qi Wang, Min Zhao. Dynamic analysis of a new aquatic ecological model based on physical and ecological integrated control[J]. Mathematical Biosciences and Engineering, 2023, 20(1): 930-954. doi: 10.3934/mbe.2023043







 DownLoad:
DownLoad: