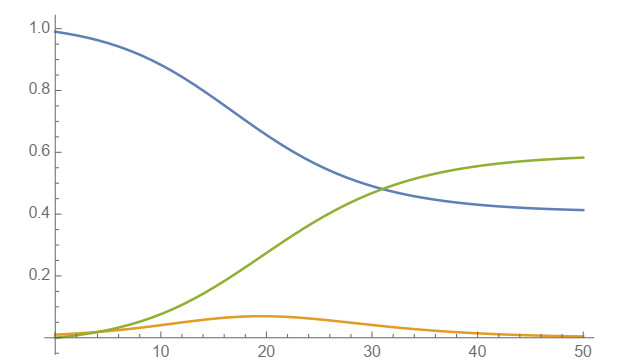
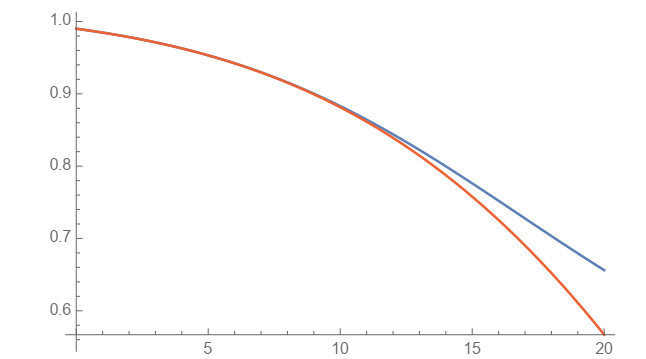
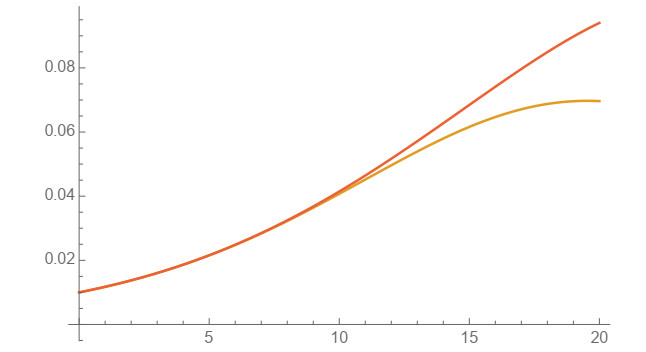
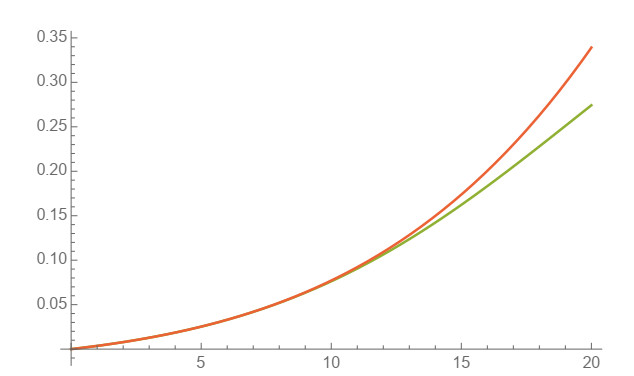
In this work, power-series solutions of compartmental epidemiological models are used to provide alternate methods to solve the corresponding systems of nonlinear differential equations. A simple and classical SIR compartmental model is considered to reveal clearly the idea of our approach. Moreover, a SAIRP compartmental model is also analyzed by using the same methodology, previously applied to the COVID-19 pandemic. Numerical experiments are performed to show the accuracy of this approach.
Citation: H. M. Srivastava, I. Area, J. J. Nieto. Power-series solution of compartmental epidemiological models[J]. Mathematical Biosciences and Engineering, 2021, 18(4): 3274-3290. doi: 10.3934/mbe.2021163
In this work, power-series solutions of compartmental epidemiological models are used to provide alternate methods to solve the corresponding systems of nonlinear differential equations. A simple and classical SIR compartmental model is considered to reveal clearly the idea of our approach. Moreover, a SAIRP compartmental model is also analyzed by using the same methodology, previously applied to the COVID-19 pandemic. Numerical experiments are performed to show the accuracy of this approach.

| [1] | F. Brauer, C. Castillo-Chavez, Z. Feng, Mathematical Models in Epidemiology, Springer-Verlag, New York, 2019. |
| [2] | W. O. Kermack, A. G. McKendrick. Contribution to the mathematical theory of epidemics, Proc. R. Soc. Lond A, 115 (1927), 700–721. |
| [3] | T. Harko, F. S. N. Lobo, M. K. Mak, Exact analytical solutions of the Susceptible-Infected-Recovered (SIR) epidemic model and of the SIR model with equal death and birth rates, Appl. Math. Comput., 236 (2014), 184–194. |
| [4] | K. Heng, C. L. Althaus, The approximately universal shapes of epidemic curves in the Susceptible-Exposed-Infectious-Recovered (SEIR) model, Sci. Rep., 10 (2020), 19365. |
| [5] | I. Area, X. H. Vidal, J. J. Nieto, M. J. P. Hermida, Determination in Galicia of the required beds at Intensive Care Units, Alexandria Eng. J., 60 (2021), 559–564. |
| [6] | F. Ndaïrou, I. Area, J. J. Nieto, D. F. M. Torres, Mathematical modeling of COVID-19 transmission dynamics with a case study of Wuhan, Chaos Solitons Fractals, 135 (2020), 109846. |
| [7] | F. Ndaïrou, I. Area, J. J. Nieto, C. Silva, D. F. M. Torres, Fractional model of COVID-19 applied to Galicia, Spain and Portugal, Chaos Solitons Fractals, 144 (2021), 110652. |
| [8] | H. M. Srivastava, Diabetes and its resulting complications: Mathematical modeling via fractional calculus, Public Health Open Access, 4 (2020), 1–5. |
| [9] | H. M. Srivastava, H. Günerhan, Analytical and approximate solutions of fractional-order susceptible-infected-recovered epidemic model of childhood disease, Math. Methods Appl. Sci., 42 (2019), 935–941. |
| [10] | H. M. Srivastava, K. M. Saad, J. F. Gómez-Aguilar, A. A. Almadiy, Some new mathematical models of the fractional-order system of human immune against IAV infection, Math. Biosci. Eng., 17 (2020), 4942–4969. |
| [11] | H. Singh, H. M. Srivastava, Z. Hammouch, K. S. Nisar, Numerical simulation and stability analysis for the fractional-order dynamics of COVID-19, Results Phys., 20 (2021), 103722. |
| [12] | Z. Liao, P. Lan, Z. Liao, Y. Zhang, S. Liu, TW-SIR: time-window based SIR for COVID-19 forecasts, Sci. Rep., 10 (2020), 22454. |
| [13] | G. D. Barmparis, G. Tsironis, Estimating the infection horizon of COVID-19 in eight countries with a data-driven approach, Chaos Solitons Fractals, 135 (2020), 09842. |
| [14] | E. B. Postnikov, Estimation of COVID-19 dynamics "on a back-of-envelope": Does the simplest SIR model provide quantitative parameters and predictions?, Chaos Solitons Fractals, 135 (2020), 109841. |
| [15] | J. Grauer, H. Löwen, B. Liebchen, Strategic spatiotemporal vaccine distribution increases the survival rate in an infectious disease like Covid-19, Sci. Rep., 10 (2020), 21594. |
| [16] | C. Hou, J. Chen, Y. Zhou, L. Hua, J. Yuan, S. He, et al., The effectiveness of quarantine of Wuhan city against the Corona Virus Disease 2019 (COVID-19): A well-mixed SEIR model analysis, J. Med. Virol., 92 (2020), 841–848. |
| [17] | Y. Zhang, X. Yu, H. Sun, G. R. Tick, W. Wei, B. Jin, Applicability of time fractional derivative models for simulating the dynamics and mitigation scenarios of COVID-19, Chaos Solitons Fractals, 138 (2020), 109959. |
| [18] | A. J. Kucharski, T. W. Russell, C. Diamond, Y. Liu, J. Edmunds, S. Funk, et al., Early dynamics of transmission and control of COVID-19: a mathematical modelling study, Lancet Infect. Dis., 20 (2020), 553–558. |
| [19] | T. Kuniya, Prediction of the Epidemic Peak of Coronavirus Disease in Japan, 2020, J. Clin. Med., 9 (2020), 789. |
| [20] | R. O. Stutt, R. Retkute, M. Bradley, G. A. Gilligan, J. Colvin, A modelling framework to assess the likely effectiveness of facemasks in combination with lock-down in managing the COVID-19 pandemic, Proc. R. Soc. A, 476 (2020), 20200376. |
| [21] | L. Zhong, L. Mu, J. Li, J. Wang, Z. Yin, D. Liu, Early Prediction of the 2019 Novel Coronavirus Outbreak in the Mainland China based on Simple Mathematical Model, IEEE Access, 2020 (2020), 19464191. |
| [22] | P. Agarwal, R. Singh, A.U. Rehman, Numerical solution of hybrid mathematical model of dengue transmission with relapse and memory via Adam-Bashforth-Moulton predictor-corrector scheme, Chaos Solitons Fractals, 143 (2021), 110564. |
| [23] | Wolfram Research, Inc., Mathematica, Version 12.2, Champaign, IL (2020). |
| [24] | C. J. Silva, C. Cruz, D. F. M. Torres, A. P. Muñuzuri, A. Carballosa, I. Area, et al., Optimal control of the COVID-19 pandemic: controlled sanitary deconfinement in Portugal, Sci. Rep., 11 (2021), 3451. |
Figures(7)

H. M. Srivastava, I. Area, J. J. Nieto. Power-series solution of compartmental epidemiological models[J]. Mathematical Biosciences and Engineering, 2021, 18(4): 3274-3290. doi: 10.3934/mbe.2021163







 DownLoad:
DownLoad: