Primary cilia are non-motile, solitary (one per cell) microtubule-based organelles that emerge from the mother centriole after cells have exited the mitotic cycle. Identified as a mechanosensing organelle that responds to both mechanical and chemical stimuli, the primary cilium provides a fertile ground for integrative investigations of mathematical modeling, numerical simulations, and experiments. Recent experimental findings revealed considerable complexity to the underlying mechanosensory mechanisms that transmit extracellular stimuli to intracellular signaling many of which include primary cilia. In this invited review, we provide a brief survey of experimental findings on primary cilia and how these results lead to various mathematical models of the mechanics of the primary cilium bent under an external forcing such as a fluid flow or a trap. Mathematical modeling of the primary cilium as a fluid-structure interaction problem highlights the importance of basal anchorage and the anisotropic moduli of the microtubules. As theoretical modeling and numerical simulations progress, along with improved state-of-the-art experiments on primary cilia, we hope that details of ciliary regulated mechano-chemical signaling dynamics in cellular physiology will be understood in the near future.
Citation: Zhangli Peng, Andrew Resnick, Y.-N. Young. Primary cilium: a paradigm for integrating mathematical modeling with experiments and numerical simulations in mechanobiology[J]. Mathematical Biosciences and Engineering, 2021, 18(2): 1215-1237. doi: 10.3934/mbe.2021066
Primary cilia are non-motile, solitary (one per cell) microtubule-based organelles that emerge from the mother centriole after cells have exited the mitotic cycle. Identified as a mechanosensing organelle that responds to both mechanical and chemical stimuli, the primary cilium provides a fertile ground for integrative investigations of mathematical modeling, numerical simulations, and experiments. Recent experimental findings revealed considerable complexity to the underlying mechanosensory mechanisms that transmit extracellular stimuli to intracellular signaling many of which include primary cilia. In this invited review, we provide a brief survey of experimental findings on primary cilia and how these results lead to various mathematical models of the mechanics of the primary cilium bent under an external forcing such as a fluid flow or a trap. Mathematical modeling of the primary cilium as a fluid-structure interaction problem highlights the importance of basal anchorage and the anisotropic moduli of the microtubules. As theoretical modeling and numerical simulations progress, along with improved state-of-the-art experiments on primary cilia, we hope that details of ciliary regulated mechano-chemical signaling dynamics in cellular physiology will be understood in the near future.

| [1] | M. Ma, M. Stoyanova, G. Rademacher, S. K. Dutcher, A. Brown, R. Zhang, Structure of the decorated ciliary doublet microtubule, Cell, 179 (2019), 909–922. |
| [2] | J. Buceta, M. Ibañes, D. Rasskin-Gutman, Y. Okada, N. Hirokawa, J. C. Izpisúa-Belmonte, Nodal cilia dynamics and the specification of the left/right axis in early vertebrate embryo development, Biophys. J., 89 (2005), 2199–2209. |
| [3] | D. Verma, F. Meng, F. Sachs, S. Z. Hua, Flow-induced focal adhesion remodeling mediated by local cytoskeletal stresses and reorganization, Cell Adhes. Migr., 9 (2015), 432–440. |
| [4] | F. Martino, A. R. Perestrelo, V. Vinarskỳ, S. Pagliari, G. Forte, Cellular mechanotransduction: from tension to function, Front. Physiol., 9 (2018), 824. |
| [5] | R. R. Ferreira, H. Fukul, R. Chow, A. Vilfan, J. Vermot, The cilium as force sensor - myth verus reality, J. Cell Sci., 132 (2019), jcs213496. |
| [6] | R. Rohatgi, W. J. Snell, The ciliary membrane, Curr. Opin. Cell Biol., 22 (2010), 541–546. |
| [7] | R. A. Bloodgood, From central to rudimentary to primary: The history of an underappreciated organelle whose time has come. the primary cilium, in Methods in Cell Biology, Vol. 94: Primary Cilia (ed. R. D. Sloboda), Academic Press, San Diego, (2009), 3–52. |
| [8] | P. Satir, L. B. Pedersen, S. T. Christensen, The primary cilium at a glance, J. Cell Sci., 123 (2010), 499. |
| [9] | H. Ishikawa, W. F. Marshall, Ciliogenesis: building the cell's antenna, Nat. Rev. Mol. Cell Biol., 12 (2011), 222–234, |
| [10] | G. J. Pazour, R. A. Bloodgood, Targeting proteins to the ciliary membrane, Curr. Top Dev. Biol., (2008), 115–149. |
| [11] | N. R. Silvester, The cilia of tetrahymena pyriformis: X-Ray diffraction by the ciliary membrane, J. Mol. Biol., 8 (1964), 11–19. |
| [12] | N. B. Gilula, P. Satir, The ciliary necklace. a ciliary membrane specialization, J. Cell Biol., 53 (1972), 494–509. |
| [13] | P. Satir, N. B. Gilula, The cell junction in a lamelibranch gill ciliated epithelium: localization of pyroantimonate precipitate, J. Cell Biol., 47 (1970), 468–487. |
| [14] | S. Temiyasathit, C. R. Jacobs, Osteocyte primary cilium and its role in bone mechanotransduction, Ann. N. Y. Acad. Sci., 1192 (2010), 422–428. |
| [15] | Z. Mirzadeh, Y.-G. Han, M. Soriano-Navarro, H. M. Garcia-Verdugo, A. Alvarez-Buylla, Cilia organize ependymal planar polarity, J. Neurosci., 30 (2010), 2600–2610. |
| [16] | C. Fisch, P. Dupuis-Williams, Ultrastructure of cilia and flagella - back to the future!, Biol. Cell, 103 (2011), 249–270. |
| [17] | D. Smith, E. Gaffney, J. Blake, Modelling mucociliary clearance, Respir. Physiol. Neurobiol., 163 (2008), 178 – 188, |
| [18] | O. E. Blacque, A. A. Sanders, Compartments within a compartment: what c. elegans can tell us about ciliary subdomain composition, biogenesis, function, and disease, Organogenesis, 10 (2014), 126–137. |
| [19] | W. M. Chong, W.-J. Wang, C.-H. Lo, T.-Y. Chiu, T.-J. Chang, Y.-P. Liu, et al., Super-resolution microscopy reveals coupling between mammalian centriole subdistal appendages and distal appendages, eLife, 9 (2020), e53580. |
| [20] | G. Garcia, J. F. Reiter, A primer on the mouse basal body, Cilia, 5 (2016), 17. |
| [21] | K. Szymanska, C. A. Johnson, The transition zone: an essential functional compartment of cilia, Cilia, 1 (2012), 10. |
| [22] | Q. Wei, K. Ling, J. Hu, The essential roles of transition fibers in the context of cilia, Curr. Opin. Cell Biol., 35 (2015), 98–105. |
| [23] | Y. Lin, P. Niewiadomski, B. Lin, H. Nakamura, S. C. Phua, J. Jiao, et al., Chemically inducible diffusion trap at cilia reveals molecular sieve-like barrier, Nat. Chem. Biol., 9 (2013), 437–443. |
| [24] | H. L. Kee, J. F. Dishinger, T. L. Blasius, C. J. Liu, B. Margolis, K. J. Verhey, A size-exclusion permeability barrier and nucleoporins characterize a ciliary pore complex that regulates transport into cilia, Nat. Cell Biol., 14 (2012), 431–437. |
| [25] | X. Jin, A. M. Mohieldin, B. S. Muntean, J. A. Green, J. V. Shah, K. Mykytyn, et al., Cilioplasm is a cellular compartment for calcium signaling in response to mechanical and chemical stimuli, Cell Mol. Life Sci., 71 (2014), 2165–2178. |
| [26] | L. Espinha, D. A. Hoey, P. R. Fernandes, H. C. Rodrigues, C. R. Jacobs, Oscillatory fluid flow influences primary cilia and microtubule mechanics, Cytoskeleton, 71 (2014), 435–445. |
| [27] | O. V. Vieira, K. Gaus, P. Verkade, J. Fullekrug, W. L. Vaz, K. Simons, Fapp2, cilium formation, and compartmentalization of the apical membrane in polarized madin-darby canine kidney (mdck) cells, Proc. Natl. Acad. Sci. USA, 103 (2006), 18556–18561. |
| [28] | P. G. DeCaen, M. Delling, T. N. Vien, D. E. Clapham, Direct recording and molecular identification of the calcium channel of primary cilia, Nature, 504 (2013), 315–318. |
| [29] | M. Delling, P. G. DeCaen, J. F. Doerner, S. Febvay, D. E. Clapham, Primary cilia are specialized calcium signalling organelles, Nature, 504 (2013), 311–314. |
| [30] | M. K. Raychowdhury, M. McLaughlin, A. J. Ramos, N. Montalbetti, R. Bouley, D. Ausiello, et al., Characterization of single channel currents from primary cilia of renal epithelial cells, J. Biol. Chem., 280 (2005), 34718–34722. |
| [31] | P. N. Inglis, K. A. Boroevich, M. R. Leroux, Piercing together a ciliome, Trends Genet., 22 (2006), 491–500. |
| [32] | D. Wloga, J. Gaertig, Post-translational modifications of microtubules, J. Cell Sci., 123 (2010), 3447–3455. |
| [33] | L. Li, X.-J. Yang, Tubulin acetylation: responsible enzymes, biological functions and human diseases, Cell. Mol. Life Sci., 72 (2015), 4237–4255. |
| [34] | C. G. Jensen, C. A. Poole, S. R. McGlashan, M. Marko, Z. I. Issa, K. V. Vujcich, et al., Ultrastructural, tomographic and confocal imaging of the chondrocyte primary cilium in situ, Cell Biol. Int., 28 (2004), 101–110. |
| [35] | I. B. Alieva, L. A. Gorgidze, Y. A. Komarova, O. A. Chernobelskaya, I. A. Vorobjev, Experimental model for studying the primary cilia in tissue culture cells, Membr. Cell Biol., 12 (1999), 895–905. |
| [36] | A. M. Mohieldin, W. A. AbouAlaiwi, M. Gao, S. M. Nauli, Chemical-fretechnique to study the ultrastructure of primary cilium, Sci. Rep., 5 (2015), 15982. |
| [37] | C. Ott, J. Lippincott-Schwartz, Visualization of live primary cilia dynamics using fluorescence microscopy, Curr. Protoc. Cell Biol., 4 (2012), 26. |
| [38] | K. E. Roth, C. L. Rieder, S. S. Bowser, Flexible-substratum technique for viewing cells from the side: some in vivo properties of primary (9+0) cilia in cultured kidney epithelia, J. Cell Sci., 89 (1998), 457–466. |
| [39] | M. Adams, U. M. Smith, C. V. Logan, C. A. Johnson, Recent advances in the molecular pathology, cell biology and genetics of ciliopathies, J. Med. Genet., 45 (2008), 257–267. |
| [40] | J. L. Badano, N. Mitsuma, P. L. Beales, N. Katsanis, The ciliopathies: an emerging class of human genetic disorders, Annu. Rev. Genomics Hum. Genet., 7 (2006), 125–148. |
| [41] | F. Hildebrandt, T. Benzing, N. Katsanis, Ciliopathies, N. Engl. J. Med., 364 (2011), 1533–1543. |
| [42] | I. Ibanez-Tallon, N. Heintz, H. Omran, To beat or not to beat: roles of cilia in development and disease, Hum. Mol. Genet., 12 (2003), R27–R35. |
| [43] | I. R. Veland, A. Awan, L. B. Pedersen, B. K. Yoder, S. T. Christensen, Primary cilia and signaling pathways in mammalian development, health and disease, Nephron. Physiol., 111 (2009), 39–53. |
| [44] | A. G. Robling, D. B. Burr, C. H. Turner, Partitionin a daily mechanical stimulus into discrete loading bouts improves the osteogenic response to loading, J. Bone Miner Res., 15 (2000), 1596–1602. |
| [45] | L. K. Saxon, A. G. Robling, A. B. Castillo, S. Mohan, C. H. Turner, The skeletal responsiveness to mechanical loading is enchanced in mice with a null mutation in estrogen receptor-beta, Am. J. Physiol. Endocrinol. Metab., 293 (2007), E484–E491. |
| [46] | T. L. Donahue, T. R. Haut, C. E. Yellowley, H. J. Donahue, C. R. Jacobs, Mechanosensitivity of bone cells to oscillating fluid flow induced shear stress may be modulated by chemotransport, J. Biomech., 36 (2003), 1363–1371. |
| [47] | K. V. der Heiden, B. P. Hierck, R. Krams, R. de Crom, C. Chen, M. Baiker, et al., Endothelial primary cilia in areas of distrubed flow are at the base of atherosclerosis, Atherosclerosis, 196 (2008), 542–550. |
| [48] | H. A. Praetorius, K. R. Spring, Bending the mdck cell primary cilium increases intracellular calcium, J. Membrane Biol., 184 (2001), 71–79. |
| [49] | H. A. Praetorius, K. R. Spring, Removal of the mdck cell primary cilium abolishes flow sensing, J. Membr. Biol., 191 (2003), 69–76. |
| [50] | S. Rydholm, G. Zwartz, J. M. Kowalewski, P. Kamali-Zare, T. Frisk, H. Brismar, Mechanical properties of primary cilia regulate the response to fluid flow, Am. J. Physiol.: Renal Physiol., 298 (2010), F1096–F1102, |
| [51] | E. J. Arnsdorf, P. Tummala, C. R. Jacobs, Wnt signaling and n-cadherin related beta-catenin signaling play a role in mechanically induced osteogenic cell fate, PLoS One, 4 (2009), e5388. |
| [52] | S. M. Nauli, F. J. Alenghat, Y. Luo, E. Williams, P. Vassilev, X. Li, et al., Polycistins 1 and 2 mediate mechanosensation in the priimary cilium of kidney cells, Nat. Genet., 33 (2003), 129–37. |
| [53] | A. I. Masyuk, T. V. Masyuk, P. L. Splinter, B. Q. Huang, A. J. Stroope, N. F. LaRusso, Cholangiocyte cilia detect changes in luminal fluid flow and transmit them into intracellular Ca2+ and cAMP signaling, Gastroenterology, 131 (2006), 911–20. |
| [54] | S. S. Pae, J. C. Saunders, Intra- and extracellular calcium modulates stereocilia stiffness on chick cochlear hair cells, Proc. Natl. Acad. Sci., 91 (1994), 1153–1157. |
| [55] | T. Y. Besschetnova, E. Kolpakova-Hart, Y. Guan, J. Zhou, B. R. Olsen, J. V. Shah, Identification of signaling pathways regulating primary cilium length, Curr. Biol., 20 (2010), 182–187. |
| [56] | E. T. OBrien, E. D. Salmon, H. P. Erickson, How calcium causes microtubule depolymerization, Cell Motil. Cytoskeleton, 36 (1997), 125–135. |
| [57] | R. H. Dillon, L. J. Fauci, An integrative model of internal axoneme mechanics and external fluid dynamics in ciliary beating, J. Theor. Biol., 207 (2000), 415–430. |
| [58] | R. H. Dillon, L. J. Fauci, C. Omoto, Mathematical modeling of axoneme mechanics and fluid dynamics in ciliary and sperm motility, Dyn. Contin. Discrete Impulsive Syst. Ser. A: Math. Anal., 10 (2003), 745–757. |
| [59] | R. H. Dillon, L. J. Fauci, C. Omoto, X. Yang, Fluid dynamic models of flagellar and ciliary beating, Ann. N. Y. Acad. Sci., 1101 (2007), 494–505. |
| [60] | R. H. Dillon, L. J. Fauci, X. Yang, Sperm motility and multicilliary beating: An integrative mechanical model, Comput. Math. Appl., 52 (2006), 749–758. |
| [61] | L. J. Fauci, R. Dillon, Biofluidmechanics of reproduction, Ann. Rev. Fluid Mechanics, 38 (2006), 371–394. |
| [62] | S. Gueron, K. Levit-Gurevich, A three-dimensional model for ciliary motion based on the internal 9+2 structure, Proc. R. Soc. Lond. B, 268 (2001), 599–607. |
| [63] | S. Gueron, K. Levit-Gurevich, The three-dimensional motion of slender filaments, Math. Methods Appl. Sci., 24 (2001), 1577–1603. |
| [64] | S. Gueron, N. Liron, Ciliary motion modeling, and dynamic multicilia interactions, Biophys. J., 63 (1992), 1045–1058. |
| [65] | M. J. Lighthill, Mathematical Biofluiddydnamics, vol. 17 of Regional Conference Series in Applied Mathematics, Society for Industrial and Applied Mathematics, 1975. |
| [66] | M. J. Lighthill, Flagellar hydrodynamics, SIAM Rev., 18 (1976), 161–230. |
| [67] | H. Shum, E. A. Gaffney, The effects of flagellar hook compliance on motility of monotrichous bacteria: A modeling study, Phys. Fluids, 24 (2012), 061901. |
| [68] | X. Yang, R. H. Dillon, L. J. Fauci, An integrative computational model of multiciliary beating, Bull. Math. Bio., 70 (2008), 1192–1215. |
| [69] | N. Coq, A. Bricard, F.-D. Delapierre, L. Malaquin, O. D. Roure, M. Fermigier, et al., Collective beating of artificial microcilia, Phys. Rev. Lett., 107 (2011), 014501. |
| [70] | A. R. Shield, B. L. Fiser, B. A. Evans, M. R. Falvo, S. Washburn, R. Superfine, Biomimetic cilia arrays generate simultaneous pumping and mixing regimes, Proc. Nat. Sci. USA, 107 (2010), 15670. |
| [71] | E. A. Schwartz, M. L. Leonard, R. Bizios, S. S. Bowser, Analysis and modeling of the primary cilium bending response to fluid shear, Am. J. Physiol.: Renal Physiol., 272 (1997), F132–F138, |
| [72] | A. Resnick, U. Hopfer, Mechanical stimulation of primary cilia, Front. Biosci., 13 (2008), 1665–1680. |
| [73] | W. Liu, N. s. Murica, Y. Duan, S. Weinbaum, B. K. Yoder, E. Schwieber, et al., Mechanoregulation of intracellular ca2+ concentration is attenuated in collecting duct of monocilium-impaired orpk mice, Am. J. Physiol. Renal Physiol., 289 (2005), F978–F988. |
| [74] | Y. N. Young, M. Downs, C. Jacobs, Dynamics of the primary cilium in shear flow, Biophys. J., 103 (2012), 629–639, |
| [75] | S. A. BABA, Flexural rigidity and elastic constant of cilia, J. Exp. Biol., 56 (1972), 459–467, |
| [76] | T. G. Fai, L. Mohapatra, P. Kar, J. Kondev, A. Amir, Length regulation of multiple flagella that self-assemble from a shared pool of components, Elife, 8 (2019), e42599. |
| [77] | E. Verghese, R. Weidenfeld, J. F. Bertram, S. D. Ricardo, J. A. Deane, Renal cilia display length alterations following tubular injury and are present early in epithelial repair, Nephrol. Dial. Transplant, 23 (2008), 834–841. |
| [78] | L. Wang, R. Weidenfeld, E. Verghese, S. D. Ricardo, J. A. Deane, J. Anat., 213 (2008), 79–85. |
| [79] | S. J. Han, H. S. Jang, J. I. Kim, J. H. Lipschutz, K. M. Park, Unilateral nephrectomy elongates primary cilia in the remaining kidney via reactive oxygen species, Sci. Rep., 2281. |
| [80] | C. G. Battle, Mechanics and Dynamics of the Primary Cilium, Ph.D thesis, 2013. |
| [81] | H. Felgner, R. Frank, M. Schliwa, Flexural rigidity of microtubules measured with the use of optical tweezers, J. Cell Sci., 109 (1996), 509–516. |
| [82] | M. Kikumoto, M. Kurachi, V. Tosa, H. Tashiro, Flexural rigidity of individual microtubules measured by a buckling force with optical traps, Biophys. J., 90 (2006), 1687–1696. |
| [83] | T. L. Hawkins, D. Sept, B. Mogessie, A. Straube, J. L. Ross, Mechanical properties of doubly stabilized microtubule filaments, Biophys. J., 104 (2013), 1517–1528. |
| [84] | Y. Gao, J. Wang, H. Gao, Persistence length of microtubules based on a continuum anisotropic shell model, J. Comput. Theor. Nanosci., 7 (2010), 1227–1237. |
| [85] | X. Liu, Y. Zhou, H. Gao, J. Wang, Anomalous flexural behaviors of microtubules, Biophys. J., 102 (2012), 1793–1803, |
| [86] | E. Memet, F. Hilitski, M. A. Morris, W. J. Schwenger, Z. Dogic, L. Mahadevan, Microtubules soften due to cross-sectional flattening, Elife, 7 (2018), e34695. |
| [87] | A. Resnick, Hif stabilization weakens primary cilia, PloS One, 11 (2016), e0165907. |
| [88] | B. Mickey, J. Howard, Rigidity of microtubules is increased by stabilizing agents., J. Cell Biol., 130 (1995), 909–917. |
| [89] | E. Verghese, J. Zhuang, D. Saiti, S. D. Ricardo, J. A. Deane, In vitro investigation of renal epithelial injury suggests that primary cilium length is regulated by hypoxia-inducible mechanisms, Cell Biol., 35 (2011), 909–913. |
| [90] | J. Flaherty, Z. Feng, Z. Peng, Y. N. Young, A. Resnick, Primary cilia have a length-dependent persistence length, Biomech. Modeling Mechanobiol., 19 (2019), 445–460, |
| [91] | J. M. Lehman, E. J. Michaud, T. R. Schoeb, Y. Aydin-Son, M. Miller, B. K. Yoder, The oak ridge polycystic kidney mouse: modeling ciliopathies of mice and men, Dev. Dyn., 237 (2008), 1960–1971. |
| [92] | J. R. Forman, S. Qamar, E. Paci, R. N. Sandford, J. Clarke, The remarkable mechanical strength of polycystin-1 supports a direct role in mechanotransduction, J. Mol. Biol., 349 (2005), 861–871. |
| [93] | F. Qian, W. Wei, G. Germino, A. Oberhauser, The nanomechanics of polycystin-1 extracellular region, J. Biol. Chem., 280 (2005), 40723–40730. |
| [94] | D. Merrick, C. A. Bertuccio, H. C. Chapin, M. Lal, V. Chauvet, M. J. Caplan, Polycystin-1 cleavage and the regulation of transcriptional pathways, Pediatric Nephrology, 29 (2014), 505–511. |
| [95] | C. M. Ta, T. N. Vien, L. C. Ng, P. G. DeCaen, Structure and function of polycystin channels in primary cilia, Cell. Signalling, 109626. |
| [96] | P. G. DeCaen, X. Liu, S. Abiria, D. E. Clapham, Atypical calcium regulation of the pkd2-l1 polycystin ion channel, Elife, 5 (2016), e13413. |
| [97] | J. R. Broekhuis, K. J. Verhey, G. Jansen, Regulation of cilium length and intraflagellar transport by the rck-kinases ick and mok in renal epithelial cells, PLoS One, 9 (2014), e108470. |
| [98] | K. Miyoshi, K. Kasahara, I. Miyazaki, M. Asanuma, Lithium treatment elongates primary cilia in the mouse brain and in cultured cells, Biochem. Biophys. Res. Commun., 388 (2009), 757–762. |
| [99] | Y. Ou, Y. Ruan, M. Chen, J. J. Moser, J. B. Rattner, F. A. van der Hoorn, Adenylate cyclase regulates elongation of mammalian primary cilia, Exp. Cell Res., 315 (2009), 2802–2817. |
| [100] | N. Sharma, Z. A. Kosan, J. E. Stallworth, N. F. Berbari, B. K. Yoder, Soluble levels of cytosolic tubulin regulate ciliary length control, Mol. Biol. Cell, 22 (2011), 806–816. |
| [101] | M. Spasic, C. R. Jacobs, Lengthening primary cilia enhances cellular mechanosensitivity, Eur. Cell Mater., 33 (2017), 158–168. |
| [102] | M. E. Downs, A. M. Nguyen, F. A. Herzog, D. A. Hoey, C. R. Jacobs, An experimental and computational analysis of primary cilia deflection under fluid flow, Comput. Methods Biomechan. Biomed. Eng., 17 (2012), 2–10, |
| [103] | C. Pozrikidis, Shear flow over cylindrical rods attached to a substrate, Fluids Struct., 26 (2010), 393–405. |
| [104] | C. Pozrikidis, Shear flow past slender elastic rods attached to a plane, Int. J.Solids Struct., 48 (2011), 137–143. |
| [105] | Y. N. Young, Dynamics of a primary cilium in time-periodic flows, Comtemporary Math., 628. |
| [106] | A. S. Popel, B. Anvari, W. E. Brownell, A. A. Spector, Modeling the mechanics of tethers pulled from the cochlear outer hair cell membrane, J. Biomechanical Eng., 130 (2008), 031007. |
| [107] | K. R. Schumacher, A. S. Popel, B. Anvari, W. E. Brownell, A. A. Spector, Computational analysis of the tether-pulling experiment to probe plasma membrane-cytoskeleton interaction in cells, Phys. Rev. E, 80 (2009), 041905. |
| [108] | Y. C. Lim, M. T. Cooling, D. S. Long, Computational models of the primary cilium and endothelial mechanotransmission, Biomech. Modeling Mechanobiol., 14 (2014), 665–678, |
| [109] | COMSOL Multiphysics Reference Manual, COMSOL Inc., Stockholm. Available from: www.comsol.com. |
| [110] | P. S. Mathieu, J. C. Bodle, E. G. Loboa, Primary cilium mechanotransduction of tensile strain in 3d culture: Finite element analyses of strain amplification caused by tensile strain applied to a primary cilium embedded in a collagen matrix, J. Biomechan., 47 (2014), 2211–2217, |
| [111] | T. J. Vaughan, C. A. Mullen, S. W. Verbruggen, L. M. McNamara, Bone cell mechanosensation of fluid flow stimulation: a fluid-structure interaction model characterising the role integrin attachments and primary cilia, Biomech. Modeling Mechanobiol., 14 (2014), 703–718, |
| [112] | H. Khayyeri, S. Barreto, D. Lacroix, Primary cilia mechanics affects cell mechanosensation: A computational study, J. Theor. Biol., 379 (2015), 38–46, |
| [113] | J. Cui, Y. Liu, B. M. Fu, Numerical study on the dynamics of primary cilium in pulsatile flows by the immersed boundary-lattice boltzmann method, Biomech. Modeling Mechanobiol., 19 (2019), 21–35, |
| [114] | Y. N. Young, L. C. Espinha, A. M. Nguyen, C. R. Jacobs, Multiscale modeling of primary cilia, in Multiscale Modeling in Biomechanics and Mechanobiology, Springer London, (2014), 87–110. |
| [115] | S. Sun, R. L. Fisher, S. S. Bowser, B. T. Pentecost, H. Sui, Three-dimensional architecture of epithelial primary cilia, Proc. Natl. Acad. Sci., 116 (2019), 9370–9379, |
| [116] | N. Stevenson, The molecular structure of primary cilia revealed by cryo-electron tomography. Available from: https://doi.org/10.1242%2Fprelights.18131. |
| [117] | Y. Wang, E. L. Botvinick, Y. Zhao, M. W. Berns, S. Usami, R. Y. Tsien, et al., Visualizing the mechanical activation of src, Nature, 434 (2005), 1040–1045, |
| [118] | Mohammad R. K. Mofrad, Roger D. Kamm, Cellular Mechanotransduction, Cambridge University Press, 2009. |
| [119] | K. Sriram, J. G. Laughlin, P. Rangamani, D. M. Tartakovsky, Shear-induced nitric oxide production by endothelial cells, Biophys. J., 84 (2003), 4087––4101, |
| [120] | E. D. Costanzo, A. I. Barakat, G. Pontrelli, Effect of flow on ATP/ADP concentration at the endothelial cell surface: interplay between shear stress and mass transport, ZAMM-J. Appl. Math. Mechan./Zeitschrift für Angewandte Math. Mechan., 98 (2018), 2222–2222. |
| [121] | B. Mazzag, A. I. Barakat, The effect of noisy flow on endothelial cell mechanotransduction: A computational study, Ann. Biomed. Eng., 39 (2010), 911–921. |
| [122] | B. M. Mazzag, J. S. Tamaresis, A. I. Barakat, A model for shear stress sensing and transmission in vascular endothelial cells, Biophys. J., 84 (2003), 4087–4101. |
| [123] | M. Beurg, J.-H. Nam, A. Crawford, R. Fettiplace, The actions of calcium on hair bundle mechanics in mammalian cochlear hair cells, Biophys. J., 94 (2008), 2639–2653. |
| [124] | S. T. Smith, R. S. Chadwick, Simulation of the response of the inner hair cell stereocilia bundle to an acoustical stimulus, PLoS ONE, 6 (2011), e18161. |
| [125] | H. Zhang, Z. Shen, B. Hogan, A. I. Barakat, C. Misbah, ATP release by red blood cells under flow: Model and simulations, Biophys. J., 115 (2018), 2218–2229. |
| [126] | R. Hinch, J. Greenstein, R. Winslow, Multi-scale models of local control of calcium induced calcium release, Progress Biophys. Mol. Biol., 90 (2006), 136–150. |
| [127] | R. Hinch, J. Greenstein, A. Tanskanen, L. Xu, R. Winslow, A simplified local control model of calcium-induced calcium release in cardiac ventricular myocytes, Biophys. J., 87 (2004), 3723–3736. |
| [128] | V. Nguyen, R. Mathias, G. Smith, A stochastic automata network descriptor for markov chain models of instantaneously coupled intracellular ca channels, Bull. Math. Biol., 67 (2005), 393–432. |
| [129] | E. A. Sobie, K. W. Dilly, J. dos Santos Cruz, W. J. Lederer, M. S. Jafri, Termination of cardiac ca2+ sparks: An investigative mathematical model of calcium-induced calcium release, Biophys. J., 83 (2002), 59–78. |
| [130] | G. S. Williams, M. A. Huertas, E. A. Sobie, M. S. Jafri, G. D. Smith, A probability density approach to modeling local control of calcium-induced calcium release in cardiac myocytes, Biophys. J., 92 (2007), 2311–2328. |
| [131] | A. Sherman, G. D. Smith, L. Dai, R. M. Miura, Asymptotic analysis of buffered calcium diffusion near a point source, SIAM J. Appl. Math., 61 (2001), 1816–1838. |
| [132] | G. Smith, Analytical steady-state solution to the rapid buffering approximation near an open ca2+ channel, Biophys. J., 71 (1996), 3064–3072. |
| [133] | M. D. Bootman, M. J. Berridge, H. Roderick, Calcium signalling: More messengers, more channels, more complexity, Curr. Biol., 12 (2002), R563–R565. |
| [134] | D. E. Clapham, Calcium signaling, Cell, 80 (1995), 259–268. |
| [135] | M. M. Maleckar, A. G. Edwards, W. E. Louch, G. T. Lines, Studying dyadic structure–function relationships: a review of current modeling approaches and new insights into ca2+ (mis)handling, Clin. Med. Insights: Cardiology, 11 (2017), 117954681769860, |
| [136] | P. A. Naik, K. R. Pardas, Finite element model to study effect of na+/k+ pump and na+/ca2+ exchanger on calcium distribution in oocytes in presence of buffers, Asian J.f Math. Stat., 7 (2014), 21–28. |
| [137] | A. Jha, N. Adlakha, Two-dimensional finite element model to study unsteady state ca2+ diffusion in neuron involving ER LEAK and SERCA, Int. J. Biomath., 08 (2015), 1550002. |
| [138] | P. A. Naik, K. R. Pardasani, Three-dimensional finite element model to study effect of RyR calcium channel, ER leak and SERCA pump on calcium distribution in oocyte cell, Int. J. Comput. Methods, 16 (2018), 1850091. |
| [139] | A. I. Barakat, Responsiveness of vascular endothelium to shear stress: potential role of ion channels and cellular cytoskeleton, Int. J. Mol. Med., 4 (1999), 323–332. |
| [140] | C. Copos, A. Mogilner, A hybrid stochastic–deterministic mechanochemical model of cell polarization, Mol. Biol. Cell, 31 (2020), 1551–1649. |
| [141] | S. S. M. Konda, J. N. Brantley, C. W. Bielawski, D. E. Makarov, Chemical reactions modulated by mechanical stress: Extended bell theory, J. Chem. Phys., 135 (2011), 164103. |
| [142] | D. H. MacLennan, W. J. Rice, N. M. Green, The mechanism of ca2+transport by sarco(endo)plasmic reticulum ca2+-ATPases, J. Biol. Chem., 272 (1997), 28815–28818. |
| [143] | M. Delling, A. A. Indzhykulian, X. Liu, Y. Li, T. Xie, D. P. Corey, D. E. Clapham, Primary cilia are not calcium-responsive mechanosensors, Nature, 531 (2016), 656–660. |
| [144] | E. H. J. Verschuren, C. Castenmiller, D. J. M. Peters, F. J. Arjona, R. J. M. Bindels, J. G. J. Hoenderop, Sensing of tubular flow and renal electrolyte transport, Nat. Rev. Nephrology, 16 (2020), 337–351. |
| [145] | G. Bossis, J. F. Brady, Self-diffusion of brownian particles in concentrated suspensions under shear, J. Chem. Phys., 87 (1987), 5437–5448. |
| [146] | A. Fletcher, J. Osborne, Seven challenges in the multiscale modelling of multicellular tissues, preprint, 2020070022. Available from: https://www.preprints.org/manuscript/202007.0022/v1. |
| [147] | E. K. Vladar, D. Antic, J. D. Axelrod, Planar cell polarity signaling: the developing cell's compass, Cold Spring Harbor Perspect. Biol., 1 (2009), a002964. |
| [148] | J. M. Reiner, Differentiation, ageing, and terminal differentiation: a semantic analysis, J. Theor. Biol., 105 (1983), 545–552. |
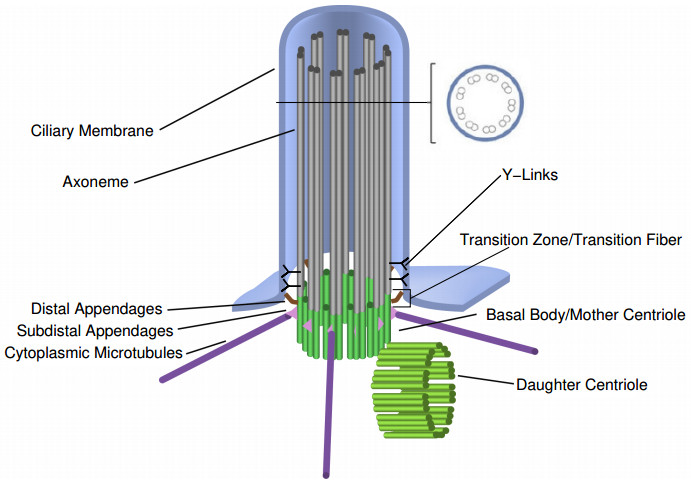
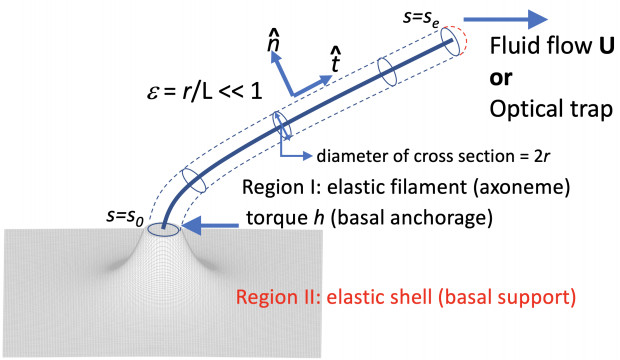
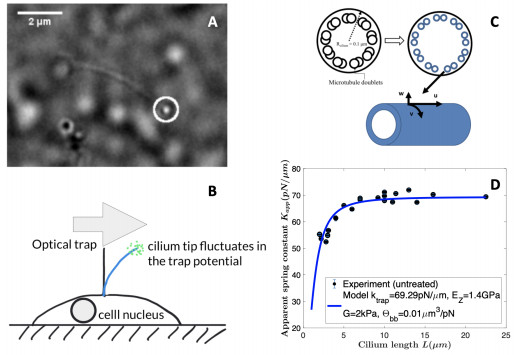
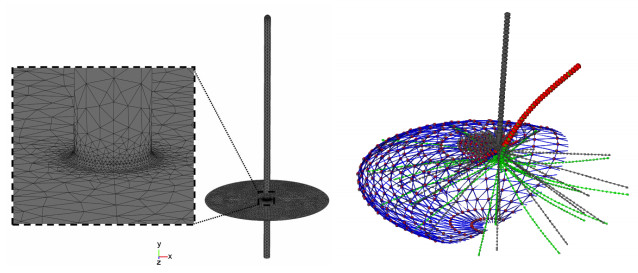
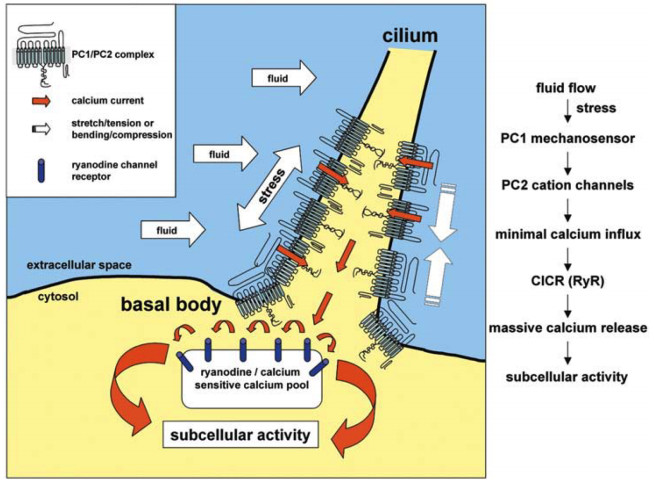
Figures(5)

Zhangli Peng, Andrew Resnick, Y.-N. Young. Primary cilium: a paradigm for integrating mathematical modeling with experiments and numerical simulations in mechanobiology[J]. Mathematical Biosciences and Engineering, 2021, 18(2): 1215-1237. doi: 10.3934/mbe.2021066







 DownLoad:
DownLoad: