Citation: Jiyu Sun, Yueming Wang, Shujun Zhang, Yunhai Ma, Jin Tong, Zhijun Zhang. The mechanism of resistance-reducing/anti-adhesion and its application on biomimetic disc furrow opener[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 4657-4677. doi: 10.3934/mbe.2020256

| [1] | D. P. Darmora, K. P. Pandey, Evaluation of performance of furrow openers of combined seed and fertiliser drills, Soil Till. Res., 34 (1995), 127-139. |
| [2] | C. Sawant, A. Kumar, I. Mani, J. K. Singh, Soil bin studies on the selection of furrow opener for conservation agriculture, J. Soil Water Conserv., 15 (2016), 107-112. |
| [3] | P. Y. Guo, M. A. Choudhary, Preliminary studies of a modified slot opener for direct drilling seeds, New Zeal. J. Exp. Agric., 1 (1985), 85-95. |
| [4] | A. Bahri, R. K. Bansal, Evaluation of different combination of furrow openers and press wheels for no till seeding, Homme. Terre. Eaux., 22 (1992), 55-66. |
| [5] | A. Ozmerzi, Seed distribution performance of furrow openers used on drill machines, AMA-Agr. Mech. Asia AF., 17 (1986), 32-34. |
| [6] | F. Ahmad, D. Weimin, D. Qishuo, M. Hussain, K. Jabran, Forces and straw cutting performance of double disc furrow opener in no-till paddy soil, PLoS One, 10 (2015), e0119648. |
| [7] | T. Vamerali, M. Bertocco, L. Sartori, Effects of a new wide-sweep opener for no-till planter on seed zone properties and root establishment in maize (Zea mays, L.): A comparison with double-disk opener, Soil Till. Res., 89 (2006), 196-209. |
| [8] | A. A. Tagar, C. Ji, J. Adamowski, J. Malard, C. S. Qi, Q. S. Ding, N. A. Abbasi, Finite element simulation of soil failure patterns under soil bin and fieldtesting conditions, Soil Till. Res., 145 (2015), 157-170. |
| [9] | H. A. Nidal, C. R. Randall, A nonlinear 3D finite element analysis of the soil forces acting on a disk plow, Soil Till. Res., 74 (2003), 115-124. |
| [10] | H. Bentaher, A. Ibrahmi, E. Hamza, M. Hbaieb, G. Kantchev, A. Maalej, W. Arnold, Finite element simulation of moldboard-soil interaction, Soil Till. Res., 134 (2013), 11-16. |
| [11] | B. A. Collins, D. B. Fowler, Effect of soil characteristics, seeding depth, operating speed, and opener design on draft force during direct seeding, Soil Till. Res., 39 (1996), 199-211. |
| [12] | S. Q. Zhang, X. Ma, C. C. Zuo, C. L. Ma, H. W. Wang, Y. J. Sun, Forces acting on disk colter and computer simulation, Trans. CSAE., 11 (1995), 52-56. |
| [13] | S. Q. Zhang, C. C. Zuo, C. L. Ma, The study on the model of disc coulter force, Trans. CSAM, 29 (1998), 71-75. |
| [14] | J. L. Jiang, L. N. Gong, M. F. Wang, Study on the working performance of the no-tillage planter unit, Trans. CSAE, 16 (2000), 64-66. |
| [15] | E. M. Tice, J. G. Hendricks, Disc coulter forces: evaluation of mathematical models, Trans. ASAE, 34 (1991), 2291-2298. |
| [16] | J. Tong, B. Z. Moayad, Y. H. Ma, J. Y. Sun, D. H. Chen, H. L. Jia, L. Q. Ren, Effects of biomimetic surface designs on furrow opener performance, J. Bionic. Eng., 6 (2009), 280-289. |
| [17] | L.Q. Ren, Progress in the bionic study on anti-adhesion and resistance-reducing of terrain machines, Sci. China Ser. E., 52 (2009), 273-284. |
| [18] | L. Q. Ren, D. X. Chen, J. G. Hu, Preliminary analysis on the rules of decreasing viscosity and removing rabbits of soil animals, J. Agr. Eng., 6 (1993), 15-20. |
| [19] | B. C. Cheng, L. Q. Ren, X. B. Xu, Bionic study on soil adhesion (two) A preliminary study on the anti-viscosity and desorption of the body surface of typical soil animals, J. Agr. Eng., 6 (1990), 2-6. |
| [20] | L. Q. Ren, Experimental Design and Optimization, Science Press, (2009). |
| [21] | D. W. Bechert, M. Bruse, W. Hage, J. G. T. van der Hoeven, G. Hoppe, Experimentson drag reducing surfaces and their optimization with an adjustable geometry, J. Fluid Mech., 338 (1997), 59-87. |
| [22] | J. B. Zhang, J. Tong, Y. H. Ma, Abrasive wear characteristics of subsoiler tines with bionic rib structure surface, Jilin Daxue Xuebao (Gongxueban),45 (2015), 174-180. |
| [23] | Z. J. Zhang, H. L. Jia, J. Y. Sun, Abrasive wear characteristics of subsoiler tines with bionic rib structure surface, Adv. Mech. Eng., 7 (2015), 1-11. |
| [24] | V. Zorba, E. Stratakis, M. Barberoglou, E. Spanakis, C. Fotakis, Biomimetic artificial surfaces quantitatively reproduce the water repellency of a lotus leaf, Adv. Mater., 20 (2008), 4049-4054. |
| [25] | H. Lee, B. P. Lee, P. B. Messersmith, A reversible wet/dry adhesive inspired by mussels and geckos, Nature, 448(2007), 338-341. |
| [26] | X. Li, T. Chen, Enhancement and suppression effects of a nanopatterned surface on bacterial adhesion, Phys. Rev. E, 93(2016), 52419. |
| [27] | D. C. Zeng, Mechanical Soil Dynamics, Beijing Science and Technology Press, (1995). |
| [28] | L. D. Bevel, W. H. Gardner, W. R. Gardner, Soil Physics, John Wiley & Sons., (1956). |
| [29] | P. G. Huray, Maxwell's Equations, Wiley-IEEE Press, (2009). |
| [30] | D. H. Qian, J. X. Zhang, A summary of study of adhesion and friction between soil and metals, Trans. CSAM, 1 (1984), 71-80. |
| [31] | D. H. Qian, A summary of study of adhesion and friction between soil and metals, Trans. CSAM, 2 (1965), 47-52. |
| [32] | L. Q. Ren, Soil Adhesion Mechanics, China Machine Press, (2011). |
| [33] | R. E. Baier, E. G. Shafrin, W. A. Zisman, Adhesion: mechanisms that assist or impede it, Science, 162 (1968), 1360-1368. |
| [34] | R. A. Fisher, Further note on the capillary forces in an ideal soil, J. Agr. Sci., 18 (1928), 406-410. |
| [35] | M. L. Nichols, The sliding of metal over soil, Agric. Eng., 6 (1925), 80-84. |
| [36] | W. B. Haines, Studies in the physical properties of soils: I. Mechanical properties concerned in cultivation, J. Agr. Sci., 15 (1925), 178-200. |
| [37] | S. Q. Deng, L. Q. Ren, Y. Liu, Z. W. Han, Tangent resistance of soil on moldboard and the mechanism of resistance reduction of bionic moldboard, J. Bionic Eng., 2 (2005), 33-46. |
| [38] | L. Q. Ren, J. Tong, J. Q. Li, B. C. Chen, Soil adhesion and biomimetics of soil-engaging components: a review, J. Agr. Eng. Res., 79 (2001), 239-263. |
| [39] | W. R. Gill, C. E. Vandenberg, Soil Dynamics in Tillage and Traction, Chinese Agricultural Machinery Press, (1983). |
| [40] | B. A. Lewis, Manual for LS-DYNA Soil Material Model 147, Federal Highway Administration, (2004). |
| [41] | I. Ahmadi, Development and assessment of a draft force calculator for disk plow using the laws of classical mechanics, Soil Till. Res.,163 (2016), 32-40. |
| [42] | A. Armin, R. Fotouhin, W. Szyszkowski, On the FE modeling of soil-blade interaction in tillage operations, Finite Elem. Anal. Des., 92 (2014), 1-11. |
| [43] | N. Eu-Gene, D. K. Aspinwall, Modeling of hard part machining, J. Mater. Process. Tech., 127 (2002), 222-229. |
| [44] | J. M. Huang, J. T. Black, An evaluation of chip separation criteria for the fem simulation of machining, J. Manuf. Sci. Eng., 118 (1996), 461-469. |
| [45] | A. P. Markopoulos, Finite Element Method in Machining Process, Springer, (2013). |
| [46] | A. Z. Shmulevich, D. Rubinstein, Interaction between soil and a wide cutting blade using the discrete element method, Soil Till. Res., 97 (2007), 37-50. |
| [47] | D. L. Jing, S. K. Yi, Electroosmotic flow in tree-like branching microchannel network, Fractals, 27 (2019), 1950095. |
| [48] | D. L. Jing, J. Song, Y. Sui, Hydraulic and thermal performances of laminar flow in fractal treelike branching microchannel network with wall velocity slip, Fractals, 28 (2020), 2050022. |
| [49] | X. Jia, Unsmooth cuticles of soil animals and theoretical analysis of their hydrophobicity and anti-soil-adhesion mechanism, J. Colloid Inter. Sci., 295 (2006), 490-494. |
| [50] | E. R. Fountaine, Investigations into the mechanism of soil adhesion, J. Soil Sci., 5 (1954), 251-263. |
| [51] | F. M. Fowkes, Additivity of intermolecular forces at interfaces. I determination of the contribution to surface and interfacial tensions of dispersion forces in various liquids, J. Phys. C, 67 (1963), 2538-2541. |
| [52] | I. Lifshitz, The kinetics of precipitation from supersaturated solid solutions, J. Phys. Chem. Solid, 19 (1961), 35-50 |
| [53] | J. Bachmann, A. Ellies, K.H. Hartgea, Development and application of a new sessile drop contact angle method to assess soil water repellency, J. Hydrol., 231-232 (2000), 66-75. |

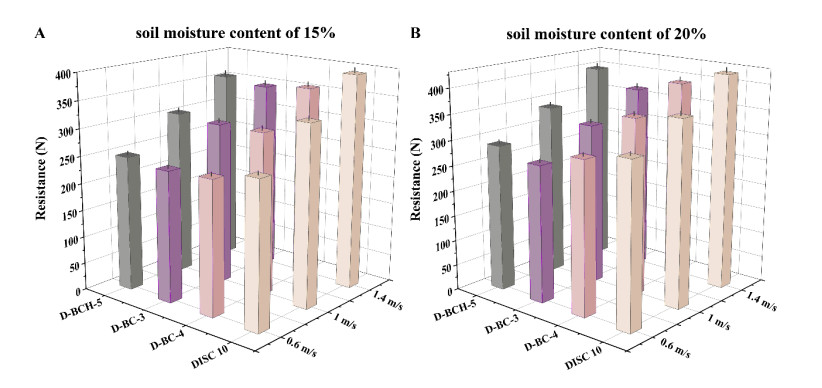
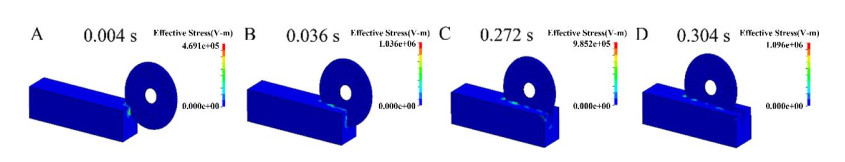
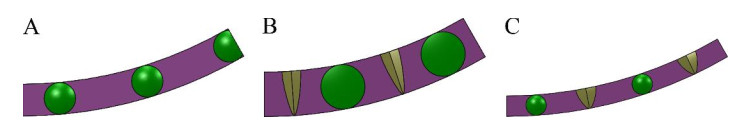
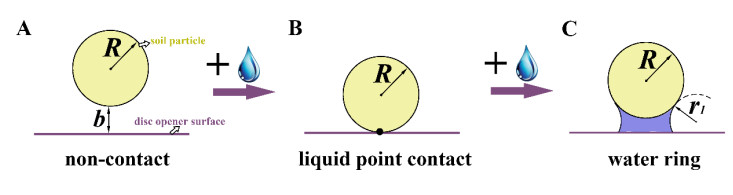
Figures(10) / Tables(6)

Jiyu Sun, Yueming Wang, Shujun Zhang, Yunhai Ma, Jin Tong, Zhijun Zhang. The mechanism of resistance-reducing/anti-adhesion and its application on biomimetic disc furrow opener[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 4657-4677. doi: 10.3934/mbe.2020256







 DownLoad:
DownLoad: