Citation: Yun-Fei Fu. Recent advances and future trends in exploring Pareto-optimal topologies and additive manufacturing oriented topology optimization[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 4631-4656. doi: 10.3934/mbe.2020255

| [1] |
K. Ghabraie, An improved soft-kill BESO algorithm for optimal distribution of single or multiple material phases, Struct. Multidiscip. Optim., 52 (2015), 773-790. doi: 10.1007/s00158-015-1268-2

|
| [2] |
K. Ghabraie, The ESO method revisited, Struct. Multidiscip. Optim., 51 (2015), 1211-1222. doi: 10.1007/s00158-014-1208-6
|
| [3] |
L. N. S. Chiu, B. Rolfe, X. H. Wu, W. Y. Yan, Effect of stiffness anisotropy on topology optimisation of additively manufactured structures, Eng. Struct., 171 (2018), 842-848. doi: 10.1016/j.engstruct.2018.05.083
|
| [4] |
O. Sigmund, K. Maute, Topology optimization approaches: A comparative review, Struct. Multidiscip. Optim., 48 (2013), 1031-1055. doi: 10.1007/s00158-013-0978-6
|
| [5] |
M. P. Bendsøe, N. Kikuchi, Generating optimal topologies in structural design using a homogenization method, Comput. Methods Appl. Mech. Eng., 71 (1988), 197-224. doi: 10.1016/0045-7825(88)90086-2
|
| [6] |
M. P. Bendsøe, Optimal shape design as a material distribution problem, Struct. Optim., 1 (1989), 193-202. doi: 10.1007/BF01650949
|
| [7] |
O. Sigmund, A 99 line topology optimization code written in Matlab, Struct. Multidiscip. Optim., 21 (2001), 120-127. doi: 10.1007/s001580050176
|
| [8] |
G. Allaire, F. Jouve, A. M. Toader, A level-set method for shape optimization, C. R. Math., 334 (2002), 1125-1130. doi: 10.1016/S1631-073X(02)02412-3
|
| [9] |
G. Allaire, F. Jouve, A. M. Toader, Structural optimization using sensitivity analysis and a level-set method, J. Comput. Phys., 194 (2004), 363-393. doi: 10.1016/j.jcp.2003.09.032
|
| [10] |
M. Y. Wang, X. M. Wang, D. M. Guo, A level set method for structural topology optimization, Comput. Methods Appl. Mech. Eng., 192 (2003), 227-246. doi: 10.1016/S0045-7825(02)00559-5
|
| [11] | Y. M. Xie, G. P. Steven, Shape and layout optimization via an evolutionary procedure, in: Proceedings of International Conference on Computational Engineering Science, Hong Kong, 1992. |
| [12] |
Y. M. Xie, G. P. Steven, A simple evolutionary procedure for structural optimization, Comput. Struct., 49 (1993), 885-896. doi: 10.1016/0045-7949(93)90035-C
|
| [13] |
X. Guo, W. S. Zhang, W. L. Zhong, Doing topology optimization explicitly and geometrically-A new moving morphable components based framework, J. Appl. Mech., 81 (2014), 081009. doi: 10.1115/1.4027609
|
| [14] |
X. Guo, W. S. Zhang, J. Zhang, J. Yuan, Explicit structural topology optimization based on moving morphable components (MMC) with curved skeletons, Comput. Methods Appl. Mech. Eng., 310 (2016), 711-748. doi: 10.1016/j.cma.2016.07.018
|
| [15] |
W. S. Zhang, J. Yuan, J. Zhang, X. Guo, A new topology optimization approach based on Moving Morphable Components (MMC) and the ersatz material model, Struct. Multidiscip. Optim., 53 (2016), 1243-1260. doi: 10.1007/s00158-015-1372-3
|
| [16] |
W. S. Zhang, D. Li, J. Yuan, J. F. Song, X. Guo, A new three-dimensional topology optimization method based on moving morphable components (MMCs), Comput. Mech., 59 (2017), 647-665. doi: 10.1007/s00466-016-1365-0
|
| [17] |
W. S. Zhang, W. Y. Yang, J. H. Zhou, D. Li, X. Guo, Structural topology optimization through explicit boundary evolution, J. Appl. Mech., 84 (2017), 011011. doi: 10.1115/1.4034972
|
| [18] |
W. S. Zhang, J. S. Chen, X. F. Zhu, J. H. Zhou, D. C. Xue, X. Lei, et al., Explicit three dimensional topology optimization via Moving Morphable Void (MMV) approach, Comput. Methods Appl. Mech. Eng., 322 (2017), 590-614. doi: 10.1016/j.cma.2017.05.002
|
| [19] |
W. S. Zhang, D. D. Li, P. Kang, X. Guo, S. K. Youn, Explicit topology optimization using IGA-based moving morphable void (MMV) approach, Comput. Methods Appl. Mech. Eng., 360 (2020), 112685. doi: 10.1016/j.cma.2019.112685
|
| [20] |
D. C. Da, L. Xia, G. Y. Li, X. D. Huang, Evolutionary topology optimization of continuum structures with smooth boundary representation, Struct. Multidiscip. Optim., 57 (2018), 2143-2159. doi: 10.1007/s00158-017-1846-6
|
| [21] |
Y. F. Fu, B. Rolfe, L. N. S. Chiu, Y. N. Wang, X. D. Huang, K. Ghabraie, Smooth topological design of 3D continuum structures using elemental volume fractions, Comput. Struct., 231 (2020), 106213. doi: 10.1016/j.compstruc.2020.106213
|
| [22] | Y. F. Fu, B. Rolfe, N. S. L. Chiu, Y. N. Wang, X. D. Huang, K. Ghabraie, SEMDOT: Smooth-Edged Material Distribution for Optimizing Topology Algorithm, arXiv preprint arXiv: 2005.09233, 2020. |
| [23] |
X. D. Huang, Smooth topological design of structures using the floating projection, Eng. Struct., 208 (2020), 110330. doi: 10.1016/j.engstruct.2020.110330
|
| [24] | X. D. Huang, On smooth or 0/1 designs of the fixed-mesh element-based topology optimization, arXiv preprint arXiv: 2006.04306 (2020). |
| [25] |
H. Bikas, P. Stavropoulos, G. Chryssolouris, Additive manufacturing methods and modelling approaches: A critical review, Int. J. Adv. Manuf. Technol., 83 (2016), 389-405. doi: 10.1007/s00170-015-7576-2
|
| [26] |
H. Lee, C. H. J. Lim, M. J. Low, N. Tham, V. M. Murukeshan, Y. J. Kim, Lasers in additive manufacturing: A review, Int. J. Precis. Eng. Manuf. Green Technol., 4 (2017), 307-322. doi: 10.1007/s40684-017-0037-7
|
| [27] |
J. C. Jiang, X. Xu, J. Stringer, Optimisation of multi-part production in additive manufacturing for reducing support waste, Virtual Phys. Prototyping, 14 (2019), 219-228. doi: 10.1080/17452759.2019.1585555
|
| [28] | J. C. Jiang, X. Xu, J. Stringer, A new support strategy for reducing waste in additive manufacturing, in: The 48th International Conference on Computers and Industrial Engineering (CIE 48), 2018. Available from: https://www.researchgate.net/publication/329999272. |
| [29] | C. W. Hull, Apparatus for production of three-dimensional objects by stereolithography, 1986. US Patent 4,575,330A, August, 1984. |
| [30] | J. C. Jiang, X. Xu, J. Stringer, Support structures for additive manufacturing: A review, J. Manuf. Mater. Process., 2 (2018), 64. |
| [31] |
J. C. Jiang, G. B. Hu, X. Li, X. Xu, P. Zheng, J. Stringer, Analysis and prediction of printable bridge length in fused deposition modelling based on back propagation neural network, Virtual Phys. Prototyping, 14 (2019), 253-266. doi: 10.1080/17452759.2019.1576010
|
| [32] |
J. C. Jiang, J. Stringer, X. Xu, Support optimization for flat features via path planning in additive manufacturing, 3D Print. Addit. Manuf., 6 (2019), 171-179. doi: 10.1089/3dp.2017.0124
|
| [33] |
J. C. Jiang, C. L. Yu, X. Xu, Y. S. Ma, J. K. Liu, Achieving better connections between deposited lines in additive manufacturing via machine learning, Math. Biosci. Eng., 17 (2020), 3382-3394. doi: 10.3934/mbe.2020191
|
| [34] |
J. C. Jiang, Y. S. Ma, Path planning strategies to optimize accuracy, quality, build time and material use in additive manufacturing: A review, Micromachines, 11 (2020), 633. doi: 10.3390/mi11070633
|
| [35] | J. C. Jiang, A novel fabrication strategy for additive manufacturing processes, J. Cleaner Production, 2020, forthcoming. |
| [36] | D. Brackett, I. Ashcroft, R. Hague, Topology optimization for additive manufacturing, in: 23rd Annual International Solid Freeform Fabrication Symposium An Additive Manufacturing Conference, Austin, TX, 2011. Available from: https://sffsymposium.engr.utexas.edu/Manuscripts/2011/2011-27-Brackett.pdf. |
| [37] |
J. K. Liu, Y. S. Ma, A survey of manufacturing oriented topology optimization methods, Adv. Eng. Software, 100 (2016), 161-175. doi: 10.1016/j.advengsoft.2016.07.017
|
| [38] |
D. Gu, W. Meiners, K. Wissenbach, R. Poprawe, Laser additive manufacturing of metallic components: Materials, processes and mechanisms, Int. Mater. Rev., 57 (2012), 133-164. doi: 10.1179/1743280411Y.0000000014
|
| [39] |
T. T. Le, S. A. Austin, S. Lim, R. A. Buswell, R. Law, A. G. Gibb, et al., Hardened properties of high-performance printing concrete, Cem. Concr. Res., 42 (2012), 558-566. doi: 10.1016/j.cemconres.2011.12.003
|
| [40] |
W. E. Frazier, Metal additive manufacturing: A review, J. Mater. Eng. Perform., 23 (2014), 1917-1928. doi: 10.1007/s11665-014-0958-z
|
| [41] |
T. Zegard, G. H. Paulino, Bridging topology optimization and additive manufacturing, Struct. Multidiscip. Optim., 53 (2016), 175-192. doi: 10.1007/s00158-015-1274-4
|
| [42] |
Y. Saadlaoui, J. L. Milan, J. M. Rossi, P. Chabrand, Topology optimization and additive manufacturing: Comparison of conception methods using industrial codes, J. Manuf. Syst., 43 (2017), 178-186. doi: 10.1016/j.jmsy.2017.03.006
|
| [43] |
A. M. Mirzendehdel, K. Suresh, Support structure constrained topology optimization for additive manufacturing, Comput. Aided Des., 81 (2016), 1-13. doi: 10.1016/j.cad.2016.08.006
|
| [44] |
H. C. Yu, J. Q. Huang, B. Zou, W. Shao, J. K. Liu, Stress-constrained shell-lattice infill structural optimisation for additive manufacturing, Virtual Phys. Prototyping, 15 (2020), 35-48. doi: 10.1080/17452759.2019.1647488
|
| [45] |
X. J. Wang, S. Q. Xu, S. W. Zhou, W. Xu, M. Leary, P. Choong, Topological design and additive manufacturing of porous metals for bone scaffolds and orthopaedic implants: A review, Biomaterials, 83 (2016), 127-141. doi: 10.1016/j.biomaterials.2016.01.012
|
| [46] |
S. Nelaturi, W. Kim, T. Kurtoglu, Manufacturability feedback and model correction for additive manufacturing, J. Manuf. Sci. Eng., 137 (2015), 021015. doi: 10.1115/1.4029374
|
| [47] | D. Thomas, The development of design rules for selective laser melting, Ph.D. thesis, University of Wales, 2009. |
| [48] |
D. Wang, Y. Q. Yang, Z. H. Yi, X. B. Su, Research on the fabricating quality optimization of the overhanging surface in SLM process, Int. J. Adv. Manuf. Technol., 65 (2013), 1471-1484. doi: 10.1007/s00170-012-4271-4
|
| [49] |
R. Mertens, S. Clijsters, K. Kempen, J. P. Kruth, Optimization of scan strategies in selective laser melting of aluminum parts with downfacing areas, J. Manuf. Sci. Eng., 136 (2014), 061012. doi: 10.1115/1.4028620
|
| [50] |
J. C. Jiang, J. Stringer, X. Xu, R. Y. Zhong, Investigation of printable threshold overhang angle in extrusion-based additive manufacturing for reducing support waste, Int. J. Comput. Integr. Manuf., 31 (2018), 961-969. doi: 10.1080/0951192X.2018.1466398
|
| [51] |
J. C. Jiang, X. Xu, J. Stringer, Optimization of process planning for reducing material waste in extrusion based additive manufacturing, Rob. Comput. Integr. Manuf., 59 (2019), 317-325. doi: 10.1016/j.rcim.2019.05.007
|
| [52] |
M. Leary, L. Merli, F. Torti, M. Mazur, M. Brandt, Optimal topology for additive manufacture: A method for enabling additive manufacture of support-free optimal structures, Mater. Des., 63 (2014), 678-690. doi: 10.1016/j.matdes.2014.06.015
|
| [53] |
X. Guo, J. H. Zhou, W. S. Zhang, Z. L. Du, C. Liu, Y. Liu, Self-supporting structure design in additive manufacturing through explicit topology optimization, Comput. Methods Appl. Mech. Eng., 323 (2017), 27-63. doi: 10.1016/j.cma.2017.05.003
|
| [54] |
R. L. Moritz, E. Reich, M. Schwarz, M. Bernt, M. Middendorf, Refined ranking relations for selection of solutions in multi objective metaheuristics, Eur. J. Oper. Res., 243 (2015), 454-464. doi: 10.1016/j.ejor.2014.10.044
|
| [55] |
J. K. Liu, A. T. Gaynor, S. K. Chen, Z. Kang, K. Suresh, A. Takezawa, et al., Current and future trends in topology optimization for additive manufacturing, Struct. Multidiscip. Optim., 57 (2018), 2457-2483. doi: 10.1007/s00158-018-1994-3
|
| [56] |
L. Meng, W. H. Zhang, D. L. Quan, G. H. Shi, L. Tang, Y. L. Hou, et al., From topology optimization design to additive manufacturing: Todays success and tomorrows roadmap, Arch. Comput. Methods Eng., 27 (2020), 805-830. doi: 10.1007/s11831-019-09331-1
|
| [57] |
L. Zadeh, Optimality and non-scalar-valued performance criteria, IEEE Trans. Autom. Control, 8 (1963), 59-60. doi: 10.1109/TAC.1963.1105511
|
| [58] | A. M. Jubril, A nonlinear weights selection in weighted sum for convex multiobjective optimization, Facta Univ., 27 (2012), 357-372. |
| [59] |
R. T. Marler, J. S. Arora, The weighted sum method for multi-objective optimization: New insights, Struct. Multidiscip. Optim., 41 (2010), 853-862. doi: 10.1007/s00158-009-0460-7
|
| [60] |
I. Turevsky, K. Suresh, Efficient generation of Pareto-optimal topologies for compliance optimization, Int. J. Numer. Methods Eng., 87 (2011), 1207-1228. doi: 10.1002/nme.3165
|
| [61] |
F. Zhao, A meshless Pareto-optimal method for topology optimization, Eng. Anal. Boundary Elem., 37 (2013), 1625-1631. doi: 10.1016/j.enganabound.2013.09.010
|
| [62] |
S. A. Gebrezgabher, M. P. Meuwissen, A. G. O. Lansink, A multiple criteria decision making approach to manure management systems in the Netherlands, Eur. J. Oper. Res., 232 (2014), 643-653. doi: 10.1016/j.ejor.2013.08.006
|
| [63] |
P. L. Yu, A class of solutions for group decision problems, Manage. Sci., 19 (1973), 841-971. doi: 10.1287/mnsc.19.8.841
|
| [64] | S. Choi, S. Kim, J. Park, S. Han, Multi-objective optimization of the inner reinforcement for a vehicles hood considering static stiffness and natural frequency, Int. J. Autom. Technol., 8 (2007), 337-342. |
| [65] |
K. H. Cho, J. Y. Park, S. P. Ryu, S. Y. Han, Reliability-based topology optimization based on bidirectional evolutionary structural optimization using multi-objective sensitivity numbers, Int. J. Autom. Technol., 12 (2011), 849-856. doi: 10.1007/s12239-011-0097-6
|
| [66] | Q. H. Zhao, X. K. Chen, L. Wang, J. F. Zhu, Z. D. Ma, Y. Lin, Simulation and experimental validation of powertrain mounting bracket design obtained from multi-objective topology optimization, Adv. Mech. Eng., 7 (2015), 1-10. |
| [67] |
K. S. Raju, P. Sonali, D. N. Kumar, Ranking of CMIP5-based global climate models for India using compromise programming, Theor. Appl. Climatol., 128 (2017), 563-574. doi: 10.1007/s00704-015-1721-6
|
| [68] |
J. Koski, R. Silvennoinen, Norm methods and partial weighting in multicriterion optimization of structures, Int. J. Numer. Methods Eng., 24 (1987), 1101-1121. doi: 10.1002/nme.1620240606
|
| [69] |
R. T. Marler, J. S. Arora, Function-transformation methods for multi-objective optimization, Eng. Optim., 37 (2005), 551-570. doi: 10.1080/03052150500114289
|
| [70] |
K. A. Proos, G. P. Steven, O. M. Querin, Y. M. Xie, Multicriterion evolutionary structural optimization using the weighting and the global criterion methods, AIAA J., 39 (2001), 2006-2012. doi: 10.2514/2.1193
|
| [71] |
I. Das, J. E. Dennis, A closer look at drawbacks of minimizing weighted sums of objectives for Pareto set generation in multicriteria optimization problems, Struct. Optim., 14, (1997), 63-69. doi: 10.1007/BF01197559
|
| [72] |
A. Messac, C. A. Mattson, Generating well-distributed sets of Pareto points for engineering design using physical programming, Optim. Eng., 3 (2002), 431-450. doi: 10.1023/A:1021179727569
|
| [73] |
K. Izui, T. Yamada, S. Nishiwaki, K. Tanaka, Multiobjective optimization using an aggregative gradient-based method, Struct. Multidiscip. Optim., 51 (2015), 173-182. doi: 10.1007/s00158-014-1125-8
|
| [74] |
Y. Sato, K. Izui, T. Yamada, S. Nishiwaki, Gradient-based multiobjective optimization using a distance constraint technique and point replacement, Eng. Optim., 48 (2016), 1226-1250. doi: 10.1080/0305215X.2015.1111068
|
| [75] |
Y. Sato, K. Izui, T. Yamada, S. Nishiwaki, Pareto frontier exploration in multiobjective topology optimization using adaptive weighting and point selection schemes, Struct. Multidiscip. Optim., 55 (2017), 409-422. doi: 10.1007/s00158-016-1499-x
|
| [76] |
Y. Sato, K. Yaji, K. Izui, T. Yamada, S. Nishiwaki, Topology optimization of a no-moving-part valve incorporating Pareto frontier exploration, Struct. Multidiscip. Optim., 56 (2017), 839-851. doi: 10.1007/s00158-017-1690-8
|
| [77] |
K. Deb, A. Pratap, S. Agarwal, T. Meyarivan, A fast and elitist multiobjective genetic algorithm: NSGA-II, IEEE Trans. Evol. Comput., 6 (2002), 182-197. doi: 10.1109/4235.996017
|
| [78] |
Y. Sato, K. Yaji, K. Izui, T. Yamada, S. Nishiwaki, An optimum design method for a thermal-fluid device incorporating multiobjective topology optimization with an adaptive weighting scheme, J. Mech. Des., 140 (2018), 031402. doi: 10.1115/1.4038209
|
| [79] |
Y. Sato, K. Izui, T. Yamada, S. Nishiwaki, Data mining based on clustering and association rule analysis for knowledge discovery in multiobjective topology optimization, Expert Syst. Appl., 119 (2019), 247-261. doi: 10.1016/j.eswa.2018.10.047
|
| [80] |
J. G. Lin, Multiple-objective problems: Pareto-optimal solutions by method of proper equality constraints, IEEE Trans. Autom. Control, 21 (1976), 641-650. doi: 10.1109/TAC.1976.1101338
|
| [81] |
K. Suresh, A 199-line Matlab code for Pareto-optimal tracing in topology optimization, Struct. Multidiscip. Optim., 42 (2010), 665-679. doi: 10.1007/s00158-010-0534-6
|
| [82] |
K. Suresh, Efficient generation of large-scale Pareto-optimal topologies, Struct. Multidiscip. Optim., 47 (2013), 49-61. doi: 10.1007/s00158-012-0807-3
|
| [83] |
S. G. Deng, K. Suresh, Multi-constrained topology optimization via the topological sensitivity, Struct. Multidiscip. Optim., 51 (2015), 987-1001. doi: 10.1007/s00158-014-1188-6
|
| [84] | A. M. Mirzendehdel, B. Rankouhi, K. Suresh, Strength-based topology optimization for anisotropic parts, Addit. Manuf., 19 (2018), 104-113. |
| [85] |
A. M. Mirzendehdel, K. Suresh, A Pareto-optimal approach to multimaterial topology optimization, J. Mech. Des., 137 (2015), 101701. doi: 10.1115/1.4031088
|
| [86] | A. Sutradhar, J. Park, P. Haghighi, J. Kresslein, D. Detwiler, J. J. Shah, Incorporating manufacturing constraints in topology optimization methods: A survey, in: ASME 2017 International Design Engineering Technical Conferences and Computers and Information in Engineering Conference, Ohio, USA, 2017. Available from: https://asmedigitalcollection.asme.org/IDETC-CIE/proceedings-abstract/IDETCCIE2017/58110/V001T02A073/259089. |
| [87] |
G. Allaire, C. Dapogny, R. Estevez, A. Faure, G. Michailidis, Structural optimization under overhang constraints imposed by additive manufacturing technologies, J. Comput. Phys., 351 (2017), 295-328. doi: 10.1016/j.jcp.2017.09.041
|
| [88] |
S. C. Subedi, C. S. Verma, K. Suresh, A review of methods for the geometric post-processing of topology optimized models, J. Comput. Inf. Sci. Eng., 20 (2020), 060801. doi: 10.1115/1.4047429
|
| [89] | P. Das, K. Mhapsekar, S. Chowdhury, R. Samant, S. Anand, Selection of build orientation for optimal support structures and minimum part errors in additive manufacturing, Comput. Aided Des. Appl., 14 (2017), 1-13. |
| [90] |
K. L. Hu, S. Jin, C. C. Wang, Support slimming for single material based additive manufacturing, Comput. Aided Des., 65 (2015), 1-10. doi: 10.1016/j.cad.2015.03.001
|
| [91] | D. S. Thomas, S. W. Gilbert, Costs and cost effectiveness of additive manufacturing, NIST Spec. Publ., 1176 (2014), 12. |
| [92] |
G. Strano, L. Hao, R. M. Everson, K. E. Evans, A new approach to the design and optimisation of support structures in additive manufacturing, Int. J. Adv. Manuf. Technol., 66 (2013), 1247-1254. doi: 10.1007/s00170-012-4403-x
|
| [93] | J. Vanek, J. A. G. Galicia, B. Benes, Clever support: Efficient support structure generation for digital fabrication, Computer Graphics Forum, 33, 2014,117-125. Available from: https://onlinelibrary.wiley.com/doi/abs/10.1111/cgf.12437. |
| [94] | J. Dumas, J. Hergel, S. Lefebvre, Bridging the gap: Automated steady scaffoldings for 3D printing, ACM Trans. Graphics, 33 (2014), 98. |
| [95] |
Y. H. Kuo, C. C. Cheng, Y. S. Lin, C. H. San, Support structure design in additive manufacturing based on topology optimization, Struct. Multidiscip. Optim., 57 (2018), 183-195. doi: 10.1007/s00158-017-1743-z
|
| [96] | F. Mezzadri, V. Bouriakov, X. P. Qian, Topology optimization of self-supporting support structures for additive manufacturing, Addit. Manuf., 21 (2018), 666-682. |
| [97] |
R. Ranjan, R. Samant, S. Anand, Integration of design for manufacturing methods with topology optimization in additive manufacturing, J. Manuf. Sci. Eng., 139 (2017), 061007. doi: 10.1115/1.4035216
|
| [98] |
K. Liu, A. Tovar, An efficient 3D topology optimization code written in Matlab, Struct. Multidiscip. Optim., 50 (2014), 1175-1196. doi: 10.1007/s00158-014-1107-x
|
| [99] | A. Panesar, M. Abdi, D. Hickman, I. Ashcroft, Strategies for functionally graded lattice structures derived using topology optimisation for additive manufacturing, Addit. Manuf., 19 (2018), 81-94. |
| [100] |
L. Cheng, A. To, Part-scale build orientation optimization for minimizing residual stress and support volume for metal additive manufacturing: Theory and experimental validation, Comput. Aided Des., 113 (2019), 1-23. doi: 10.1016/j.cad.2019.03.004
|
| [101] | L. Cheng, X. Liang, J. X. Bai, Q. Chen, J. Lemon, A. To, On utilizing topology optimization to design support structure to prevent residual stress induced build failure in laser powder bed metal additive manufacturing, Addit. Manuf., 27 (2019), 290-304. |
| [102] | A. T. Gaynor, N. A. Meisel, C. B. Williams, J. K. Guest, Topology optimization for additive manufacturing: Considering maximum overhang constraint, in: 15th AIAA/ISSMO Multidisciplinary Analysis and Optimization Conference, Atlanta, GA, 2014. Available from: https://arc.aiaa.org/doi/10.2514/6.2014-2036. |
| [103] |
A. T. Gaynor, J. K. Guest, Topology optimization considering overhang constraints: Eliminating sacrificial support material in additive manufacturing through design, Struct. Multidiscip. Optim., 54 (2016), 1157-1172. doi: 10.1007/s00158-016-1551-x
|
| [104] | M. Langelaar, Topology optimization of 3D self-supporting structures for additive manufacturing, Addit. Manuf., 12 (2016) 60-70. |
| [105] |
M. Langelaar, An additive manufacturing filter for topology optimization of print-ready designs, Struct. Multidiscip. Optim., 55 (2017), 871-883. doi: 10.1007/s00158-016-1522-2
|
| [106] | B. Barroqueiro, A. Andrade-Campos, R. A. F. Valente, Designing self supported SLM structures via topology optimization, J. Manuf. Mater. Process., 3 (2019), 68. |
| [107] |
Y. F. Fu, B. Rolfe, N. S. L. Chiu, Y. N. Wang, X. D. Huang, K. Ghabraie, Design and experimental validation of self-supporting topologies for additive manufacturing, Virtual Phys. Prototyping, 14 (2019), 382-394. doi: 10.1080/17452759.2019.1637023
|
| [108] |
Y. F. Fu, B. Rolfe, N. S. L. Chiu, Y. N. Wang, X. D. Huang, K. Ghabraie, Parametric studies and manufacturability experiments on smooth self-supporting topologies, Virtual Phys. Prototyping, 15 (2020), 22-34. doi: 10.1080/17452759.2019.1644185
|
| [109] | E. van de Ven, R. Maas, C. Ayas, M. Langelaar, F. van Keulen, Continuous front propagation-based overhang control for topology optimization with additive manufacturing, Struct. Multidiscip. Optim., 57 (2018), 2075-2091. |
| [110] | A. G. Jimenez, R. A. Loyola, J. Santamaría, I. F. de Bustos, A new overhang constraint for topology optimization of self-supporting structures in additive manufacturing, Struct. Multidiscip. Optim., 58 (2018), 2003-2017. |
| [111] |
A. Garaigordobil, R. Ansola, E. Vegueria, I. Fernandez, Overhang constraint for topology optimization of self-supported compliant mechanisms considering additive manufacturing, Comput. Aided Des., 109 (2019), 33-48. doi: 10.1016/j.cad.2018.12.006
|
| [112] | Y. H. Kuo, C. C. Cheng, Self-supporting structure design for additive manufacturing by using a logistic aggregate function, Struct. Multidiscip. Optim., 2019 (2019), 1-13. |
| [113] |
K. Q. Zhang, G. D. Cheng, L. Xu, Topology optimization considering overhang constraint in additive manufacturing, Comput. Struct., 212 (2019), 86-100. doi: 10.1016/j.compstruc.2018.10.011
|
| [114] |
G. Allaire, C. Dapogny, R. Estevez, A. Faure, G. Michailidis, Structural optimization under overhang constraints imposed by additive manufacturing processes: An overview of some recent results, Appl. Math. Nonlinear Sci., 2 (2017), 385-402. doi: 10.21042/AMNS.2017.2.00031
|
| [115] | G. Allaire, C. Dapogny, A. Faure, G. Michailidis, A model of layer by layer mechanical constraint for additive manufacturing in shape and topology optimization, in: 12th World Congress on Structural and Multidisciplinary Optimization (WCSMO12), Brunswick, Germany, 2017. Available from: https://hal.archives-ouvertes.fr/hal-01536668/. |
| [116] |
G. Allaire, C. Dapogny, A. Faure, G. Michailidis, Shape optimization of a layer by layer mechanical constraint for additive manufacturing, C. R. Math., 355 (2017), 699-717. doi: 10.1016/j.crma.2017.04.008
|
| [117] |
J. K. Liu, A. C. To, Deposition path planning-integrated structural topology optimization for 3D additive manufacturing subject to self-support constraint, Comput. Aided Des., 91 (2017), 27-45. doi: 10.1016/j.cad.2017.05.003
|
| [118] |
J. K. Liu, Y. S. Ma, A. J. Qureshi, R. Ahmad, Light-weight shape and topology optimization with hybrid deposition path planning for FDM parts, Int. J. Adv. Manuf. Technol., 97 (2018), 1123-1135. doi: 10.1007/s00170-018-1955-4
|
| [119] |
W. H. Zhang, L. Zhou, Topology optimization of self-supporting structures with polygon features for additive manufacturing, Comput. Methods Appl. Mech. Eng., 334 (2018), 56-78. doi: 10.1016/j.cma.2018.01.037
|
| [120] |
J. Q. Huang, Q. Chen, H. Jiang, B. Zou, L. Li, J. K. Liu, et al., A survey of design methods for material extrusion polymer 3d printing, Virtual Phys. Prototyping, 15 (2020), 148-162. doi: 10.1080/17452759.2019.1708027
|
| [121] |
J. Liu, H. Yu, Self-support topology optimization with horizontal overhangs for additive manufacturing, J. Manuf. Sci. Eng., 142 (2020), 091003. doi: 10.1115/1.4047352
|
| [122] | Y. Mass, O. Amir, Topology optimization for additive manufacturing: Accounting for overhang limitations using a virtual skeleton, Addit. Manuf., 18 (2017), 58-73. |
| [123] | A. M. Driessen, Overhang constraint in topology optimisation for additive manufacturing: A density gradient based approach, Master's thesis, Delft University of Technology, 2016. |
| [124] | M. Langelaar, Combined optimization of part topology, support structure layout and build orientation for additive manufacturing, Struct. Multidiscip. Optim., 57 (2018), 1985-2004. |
| [125] |
J. K. Guest, J. H. Prévost, T. Belytschko, Achieving minimum length scale in topology optimization using nodal design variables and projection functions, Int. J. Numer. Methods Eng., 61 (2004), 238-254. doi: 10.1002/nme.1064
|
| [126] |
J. K. Guest, Imposing maximum length scale in topology optimization, Struct. Multidiscip. Optim., 37 (2009), 463-473. doi: 10.1007/s00158-008-0250-7
|
| [127] |
J. K. Guest, Topology optimization with multiple phase projection, Comput. Methods Appl. Mech. Eng., 199 (2009), 123-135. doi: 10.1016/j.cma.2009.09.023
|
| [128] |
W. S. Zhang, W. L. Zhong, X. Guo, An explicit length scale control approach in SIMP-based topology optimization, Comput. Methods Appl. Mech. Eng., 282 (2014), 71-86. doi: 10.1016/j.cma.2014.08.027
|
| [129] |
M. D. Zhou, B. S. Lazarov, F. W. Wang, O. Sigmund, Minimum length scale in topology optimization by geometric constraints, Comput. Methods Appl. Mech. Eng., 293 (2015), 266-282. doi: 10.1016/j.cma.2015.05.003
|
| [130] |
W. S. Zhang, D. Li, J. Zhang, X. Guo, Minimum length scale control in structural topology optimization based on the Moving Morphable Components (MMC) approach, Comput. Methods Appl. Mech. Eng., 311 (2016), 327-355. doi: 10.1016/j.cma.2016.08.022
|
| [131] |
G. Allaire, F. Jouve, G. Michailidis, Thickness control in structural optimization via a level set method, Struct. Multidiscip. Optim., 53 (2016), 1349-1382. doi: 10.1007/s00158-016-1453-y
|
| [132] |
L. Hägg, E. Wadbro, On minimum length scale control in density based topology optimization, Struct. Multidiscip. Optim., 58 (2018), 1015-1032. doi: 10.1007/s00158-018-1944-0
|
| [133] |
X. P. Qian, Undercut and overhang angle control in topology optimization: A density gradient based integral approach, Int. J. Numer. Methods Eng., 111 (2017), 247-272. doi: 10.1002/nme.5461
|
| [134] |
S. R. Mohan, S. Simhambhatla, Adopting feature resolution and material distribution constraints into topology optimisation of additive manufacturing components, Virtual Phys. Prototyping, 14 (2019), 79-91. doi: 10.1080/17452759.2018.1501275
|
| [135] |
J. K. Liu, Piecewise length scale control for topology optimization with an irregular design domain, Comput. Methods Appl. Mech. Eng., 351 (2019), 744-765. doi: 10.1016/j.cma.2019.04.014
|
| [136] |
J. K. Liu, Y. F. Zheng, R. Ahmad, J. Y. Tang, Y. S. Ma, Minimum length scale constraints in multiscale topology optimisation for additive manufacturing, Virtual Phys. Prototyping, 14 (2019) 229-241. doi: 10.1080/17452759.2019.1584944
|
| [137] |
S. T. Liu, Q. H. Li, W. J. Chen, L. Y. Tong, G. D. Cheng, An identification method for enclosed voids restriction in manufacturability design for additive manufacturing structures, Front. Mech. Eng., 10 (2015), 126-137. doi: 10.1007/s11465-015-0340-3
|
| [138] |
Q. H. Li, W. J. Chen, S. T. Liu, H. R. Fan, Topology optimization design of cast parts based on virtual temperature method, Comput. Aided Des., 94 (2018), 28-40. doi: 10.1016/j.cad.2017.08.002
|
| [139] |
L. Zhou, W. H. Zhang, Topology optimization method with elimination of enclosed voids, Struct. Multidiscip. Optim., 60 (2019), 117-136. doi: 10.1007/s00158-019-02204-y
|
| [140] | Y. L. Xiong, S. Yao, Z. L. Zhao, Y. M. Xie, A new approach to eliminating enclosed voids in topology optimization for additive manufacturing, Addit. Manuf., 32 (2020), 101006. |
| [141] | K. Suresh, A. Ramani, A. Kaushik, An adaptive weighting strategy for multi-load topology optimization, in: ASME 2012 International Design Engineering Technical Conferences and Computers and Information in Engineering Conference, Chicago, USA, 2012. Available from: https://asmedigitalcollection.asme.org/IDETC-CIE/proceedings-abstract/IDETCCIE2012/45011/1295/254817. |
| [142] |
H. Li, L. Gao, P. G. Li, Topology optimization of structures under multiple loading cases with a new compliance-volume product, Eng. Optim., 46 (2014), 725-744. doi: 10.1080/0305215X.2013.800054
|
| [143] |
M. Baumers, L. Beltrametti, A. Gasparre, R. Hague, Informing additive manufacturing technology adoption: Total cost and the impact of capacity utilisation, Int. J. Prod. Res., 55 (2017), 6957-6970. doi: 10.1080/00207543.2017.1334978
|
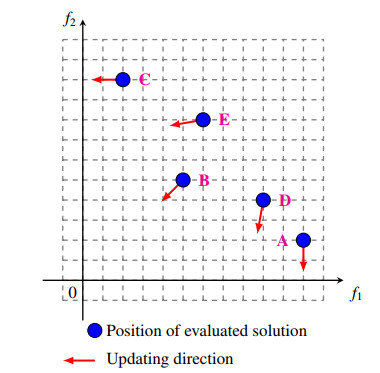
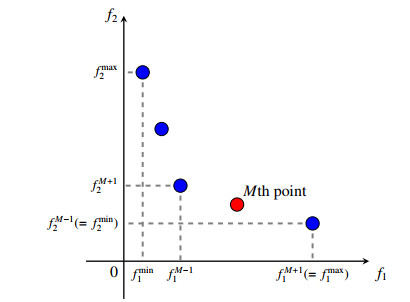
Figures(2)

Yun-Fei Fu. Recent advances and future trends in exploring Pareto-optimal topologies and additive manufacturing oriented topology optimization[J]. Mathematical Biosciences and Engineering, 2020, 17(5): 4631-4656. doi: 10.3934/mbe.2020255







 DownLoad:
DownLoad: