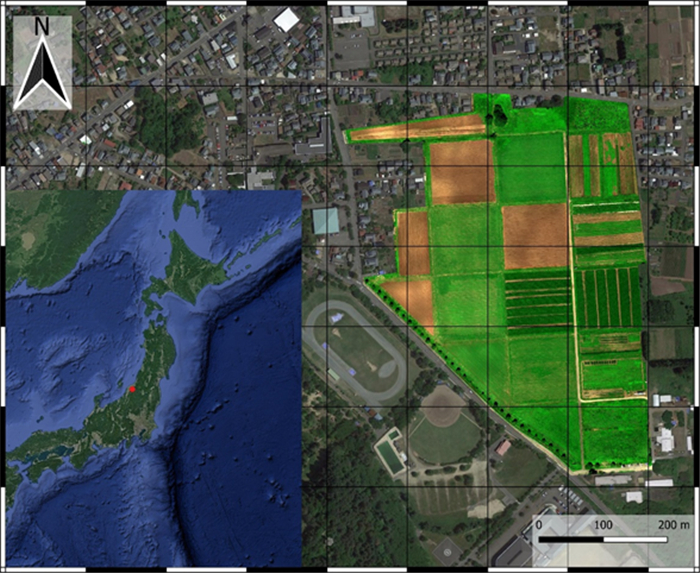
Agriculture's sustainability is a subject of concern, and digital farming technology has been proposed to solve this problem. The digital revolution is transforming agriculture by utilizing modern equipment, computerized tools, and information and communication technology (ICT) to improve decision-making and productivity. Digital farming technology enables even inexperienced farmers to apply such techniques by using the IoT and AI to collect and analyze data from their farming practices and the surrounding environment to improve productivity. The versatile mapping platform Mapbox is utilized to construct the map, which allows for custom data layer integration and interactive features. Backend development is performed using the Java programming language, which facilitates seamless data processing, storage, and retrieval. The interactive map allows for dynamic overlays of crucial information, such as plot numbers, measurements, crop details, crop health assessments, NDVI, RGB, and DEM. The study involved data collection, analysis of the data, and thematic layer development using GIS to create interactive maps. In this research, two sets of DJI drones, Agisoft Metashape software, QGIS, and Mapbox were used to collect and prepare the data for the interactive map. The data was used to create the results, which were web maps that had several interactive features, such as "display popup on hover, " "swipe between maps, " and "change a map's style." The result was a thematic layer of information such as RGB, NDVI, DEM, and other field information. This research demonstrated the benefits and applicability of information technology for digital transformation in agriculture under the DX Project launched at Niigata University, Japan. This aids in the goal of producing interactive agricultural maps based on map classification, content element analysis, the development of GIS capabilities, and remote sensing data.
Citation: Ayomikun D. Ajayi, Boris Boiarskii, Kouya Aoyagi, Hideo Hasegawa. Utilizing MapBox API, Java, and ICT in the creation of agricultural interactive maps for improved farm management and decision-making[J]. AIMS Agriculture and Food, 2024, 9(2): 393-410. doi: 10.3934/agrfood.2024023
Agriculture's sustainability is a subject of concern, and digital farming technology has been proposed to solve this problem. The digital revolution is transforming agriculture by utilizing modern equipment, computerized tools, and information and communication technology (ICT) to improve decision-making and productivity. Digital farming technology enables even inexperienced farmers to apply such techniques by using the IoT and AI to collect and analyze data from their farming practices and the surrounding environment to improve productivity. The versatile mapping platform Mapbox is utilized to construct the map, which allows for custom data layer integration and interactive features. Backend development is performed using the Java programming language, which facilitates seamless data processing, storage, and retrieval. The interactive map allows for dynamic overlays of crucial information, such as plot numbers, measurements, crop details, crop health assessments, NDVI, RGB, and DEM. The study involved data collection, analysis of the data, and thematic layer development using GIS to create interactive maps. In this research, two sets of DJI drones, Agisoft Metashape software, QGIS, and Mapbox were used to collect and prepare the data for the interactive map. The data was used to create the results, which were web maps that had several interactive features, such as "display popup on hover, " "swipe between maps, " and "change a map's style." The result was a thematic layer of information such as RGB, NDVI, DEM, and other field information. This research demonstrated the benefits and applicability of information technology for digital transformation in agriculture under the DX Project launched at Niigata University, Japan. This aids in the goal of producing interactive agricultural maps based on map classification, content element analysis, the development of GIS capabilities, and remote sensing data.

| [1] |
Turnbull D (1996) Cartography and science in early modern Europe: Mapping the construction of knowledge spaces. Imago Mundi 48: 5–24. https://doi.org/10.1080/03085699608592830 doi: 10.1080/03085699608592830

|
| [2] |
Borges LF, Bazzi CL, de Souza EG, et al. (2020) Web software to create thematic maps for precision agriculture. Pesq Agropec Bras 55: 00735. https://doi.org/10.1590/S1678-3921.PAB2020.V55.00735 doi: 10.1590/S1678-3921.PAB2020.V55.00735
|
| [3] | Whelan B, Taylor J (2019) Precision Agriculture for Grain Production Systems, CSIRO Publishing. https://doi.org/10.1071/9780643107489 |
| [4] |
Bellón B, Bégué A, Lo Seen D, et al. (2017) A remote sensing approach for regional-scale mapping of agricultural land-use systems based on NDVI time series. Remote Sens 9: 600. https://doi.org/10.3390/rs9060600 doi: 10.3390/rs9060600
|
| [5] |
Bronson K (2019) Looking through a responsible innovation lens at uneven engagements with digital farming. NJAS-Wageningen J Life Sci 90–91: 1–6. https://doi.org/10.1016/j.njas.2019.03.001 doi: 10.1016/j.njas.2019.03.001
|
| [6] |
Smith DA (2016) Online interactive thematic mapping: Applications and techniques for socio-economic research. Comput, Environ Urban Syst 57: 106–117. https://doi.org/10.1016/j.compenvurbsys.2016.01.002 doi: 10.1016/j.compenvurbsys.2016.01.002
|
| [7] |
Oymatov R, Safayev S (2021) Creation of a complex electronic map of agriculture and agro-geo databases using GIS techniques. E3S Web Conf 258: 03020. https://doi.org/10.1051/e3sconf/202125803020 doi: 10.1051/e3sconf/202125803020
|
| [8] |
Oymatov RK, Mamatkulov ZJ, Reimov MP, et al. (2021) Methodology development for creating agricultural interactive maps. IOP Conf Ser: Earth Environ Sci 868: 012074. https://doi.org/10.1088/1755-1315/868/1/012074 doi: 10.1088/1755-1315/868/1/012074
|
| [9] |
Degbelo A, Kruse J, Pfeiffer M (2019) Interactive maps, productivity and user experience: A user study in the e-mobility domain. Trans GIS 23: 1352–1373. https://doi.org/10.1111/tgis.12576 doi: 10.1111/tgis.12576
|
| [10] |
Kamel Boulos MN (2003) The use of interactive graphical maps for browsing medical/health Internet information resources. Int J Health Geographics 2: 1. https://doi.org/10.1186/1476-072X-2-1 doi: 10.1186/1476-072X-2-1
|
| [11] |
Roth RE (2013) Interactive maps: What we know and what we need to know. J Spat Inf Sci 6: 59–115. https://doi.org/10.5311/JOSIS.2013.6.105 doi: 10.5311/JOSIS.2013.6.105
|
| [12] |
Haklay M, Singleton A, Parker C (2008) Web mapping 2.0: The neogeography of the GeoWeb. Geogr Compass 2: 2011–2039. https://doi.org/10.1111/j.1749-8198.2008.00167.x doi: 10.1111/j.1749-8198.2008.00167.x
|
| [13] | Peterson MP (2020) Map Interactivity. In: Kobayashi A (Ed.), International Encyclopedia of Human Geography, 2 ed., 259–262. https://doi.org/10.1016/B978-0-08-102295-5.10566-9 |
| [14] |
Horbiński T, Lorek D (2020) The use of Leaflet and GeoJSON files for creating the interactive web map of the preindustrial state of the natural environment. J Spat Sci 67: 61–77. https://doi.org/10.1080/14498596.2020.1713237 doi: 10.1080/14498596.2020.1713237
|
| [15] |
Medyńska-Gulij B (2012) Pragmatische Kartographie in Google Maps API. KN-J Cartography Geogr Inf 62: 250–255. https://doi.org/10.1007/BF03544493 doi: 10.1007/BF03544493
|
| [16] | Wagaskar K, Joshi N, Tripathy AK, et al. (2018) Intelligent AgriTrade to Abet Indian Farming. In: Abraham A, Muhuri PK, Muda AK (Eds.), Advances in Intelligent Systems and Computing, 736: 932–941. https://doi.org/10.1007/978-3-319-76348-4_89 |
| [17] |
Lin YP, Petway JR, Anthony J, et al. (2017) Blockchain: The evolutionary next step for ICT e-agriculture. Environments 4: 50. https://doi.org/10.3390/environments4030050 doi: 10.3390/environments4030050
|
| [18] |
Zubarev Y, Fomin D, Zubarev N (2019) Using high-precision farming systems in the agricultural sector - the path to digital agriculture. Adv Intell Syst Res 167: 104–107. https://doi.org/10.2991/ispc-19.2019.23 doi: 10.2991/ispc-19.2019.23
|
| [19] |
Boiarskii B, Hasegawa H (2019) Comparison of NDVI and NDRE indices to detect differences in vegetation and chlorophyll content. J Mech Contin Math Sci 4: 20–29. https://doi.org/10.26782/jmcms.spl.4/2019.11.00003 doi: 10.26782/jmcms.spl.4/2019.11.00003
|
| [20] | Velkoski G, Gusev M, Ristov S (2017) Analysis of interactive image technologies. In: 2016 24th Telecommunications Forum (TELFOR), Belgrade, Serbia, 1–4. https://doi.org/10.1109/TELFOR.2016.7818929 |
| [21] |
Peterson MP (2021) A comparison of feature density for large scale online maps. Cartographic Perspect 97: 26–42. https://doi.org/10.14714/CP97.1707 doi: 10.14714/CP97.1707
|
| [22] | API Reference | Mapbox GL JS | Mapbox. Available from: https://docs.mapbox.com/mapbox-gl-js/api/. |
| [23] | Dorman M (2020) Introduction to Web Mapping, 1st ed, New York. Available from: https://doi.org/10.1201/9780429352874 |
| [24] |
Doernhoefer M (2006) Surfing the net for software engineering notes. ACM SIGSOFT Software Eng Notes 31: 5–13. https://doi.org/10.1145/1108768.1108783 doi: 10.1145/1108768.1108783
|
| [25] |
Costanza P, Herzeel C, Verachtert W (2019) A comparison of three programming languages for a full-fledged next-generation sequencing tool. BMC Bioinf 20: 301. https://doi.org/10.1186/s12859-019-2903-5 doi: 10.1186/s12859-019-2903-5
|
| [26] | Leavesley GH, David O, Garen D, et al. (2008) A modeling framework for improved agricultural water-supply forecasting. AGU Fall Meet 2008: C21A–0497. |
| [27] |
Latif S, Beck F (2019) Interactive map reports summarizing bivariate geographic data. Visual Inf 3: 27–37. https://doi.org/10.1016/J.VISINF.2019.03.004 doi: 10.1016/J.VISINF.2019.03.004
|
| [28] |
Maiellaro N, Varasano A (2017) One-page multimedia interactive map. ISPRS Int J Geo-Inf 6: 34. https://doi.org/10.3390/ijgi6020034 doi: 10.3390/ijgi6020034
|
| [29] |
Di Pasquale D, Lerario A, Maiellaro N, et al. (2013) Open source interactive map of Albania cultural heritage. IERI Proc 4: 383–390. https://doi.org/10.1016/J.IERI.2013.11.055 doi: 10.1016/J.IERI.2013.11.055
|
| [30] |
She J, Liu J, Li C, et al. (2017) A line-feature label placement algorithm for interactive 3D map. Comput Graphics 67: 86–94. https://doi.org/10.1016/J.CAG.2017.06.002 doi: 10.1016/J.CAG.2017.06.002
|
| [31] |
Oymatov R, Mamatkulov Z, Maksudov R, et al. (2023) Improving the methods of agricultural mapping using remote sensing data. E3S Web Conf 386: 04008. https://doi.org/10.1051/e3sconf/202338604008 doi: 10.1051/e3sconf/202338604008
|
| [32] |
Kumalawati R, Salamiah, Yuliarti A, et al. (2021) Potential mapping agricultural commodities to mitigation of food problem in the future. Geojournal Tourism Geosites 33: 1480–1485. https://doi.org/10.30892/gtg.334spl05-596 doi: 10.30892/gtg.334spl05-596
|
| [33] |
Nurdin DP, Karim S, Kurniadin N (2022) Geographic information system for mapping agricultural land in North Samarinda district. TEPIAN 3: 165–172. https://doi.org/10.51967/tepian.v3i3.701 doi: 10.51967/tepian.v3i3.701
|
| [34] |
Stojanovic V, Falconer RE, Isaacs J, et al. (2017) Streaming and 3D mapping of AGRI-data on mobile devices. Comput Electron Agric 138: 188–199. https://doi.org/10.1016/j.compag.2017.03.019 doi: 10.1016/j.compag.2017.03.019
|
| [35] | Rzeszewski M (2023) Mapbox. In: Burnett CM (Ed.), Evaluating Participatory Mapping Software, Springer International Publishing, Cham, 21–40. https://doi.org/10.1007/978-3-031-19594-5_2 |
| [36] |
Den Besten N, Steele-Dunne S, de Jeu R, et al. (2021) Towards monitoring waterlogging with remote sensing for sustainable irrigated agriculture. Remote Sens 13: 2929. https://doi.org/10.3390/rs13152929 doi: 10.3390/rs13152929
|
| [37] | Boiarskii B, Sinegovskii M (2022) Application of NDVI and NDRE vegetation indices in the assessment of soybean productivity under nitrogen controlled-release fertilizer. 2022 8th International Conference on Information Technology and Nanotechnology, ITNT, IEEE, 1–6. https://doi.org/10.1109/ITNT55410.2022.9848588 |
| [38] | Murray JD, Van Ryper W (1997) Encyclopedia of graphics file formats, 2nd Ed., Sebastopol: O'Reilly & Associates Inc. |
| [39] |
Tucker CJ (1979) Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens Environ 8: 127–150. https://doi.org/10.1016/0034-4257(79)90013-0 doi: 10.1016/0034-4257(79)90013-0
|
| [40] | Cadenas C (2014) Geovisualization: Integration and visualization of multiple datasets using Mapbox. Computer Engineering. |
| [41] | Kennedy B, Musciano C (2002) HTML & XHTML—The Definitive Guide, 5th Ed., Sebastopol: O'Reilly Media Inc. |
| [42] | Wang W, Poo-Caamaño G, Wilde E, et al. (2015) What is the GIST? Understanding the use of public gists on GitHub. IEEE International Working Conference on Mining Software Repositories. https://doi.org/10.1109/MSR.2015.36 |
| [43] |
Netek, Brus, Tomecka (2019) Performance testing on marker clustering and heatmap visualization techniques: A comparative study on JavaScript mapping libraries. ISPRS Int J Geo-Inf 8: 348. https://doi.org/10.3390/ijgi8080348 doi: 10.3390/ijgi8080348
|
| [44] | Giriprasad Patil, Girish MK, Mrs. Vidya (2022) A review paper on new web development technology. Int J Adv Res Sci, Commun Technol 2: 468–476. https://doi.org/10.48175/ijarsct-2901 |
| [45] | Shahu Gaikwad S, Adkar P (2019) A review paper on bootstrap framework. IRE J 2: 349–351. |
| [46] | Arapov P, Buffa M, ben Othmane A (2014) Developing web of data applications from the browser. WWW 2014 Companion-Proceedings of the 23rd International Conference on World Wide Web, 599–602. https://doi.org/10.1145/2567948.2578040 |
| [47] |
Hernando R, Macías JA (2023) Development of usable applications featuring QR codes for enhancing interaction and acceptance: A case study. Behav Inf Technol 42: 360–378. https://doi.org/10.1080/0144929X.2021.2022209 doi: 10.1080/0144929X.2021.2022209
|
| [48] | Deineko Z, Kraievska N, Lyashenko V (2022) QR code as an element of educational activity. Int J Acad Inf Syst Res 6: 26–31. |
| [49] | Marín JF, Parra L, Lloret J, et al. (2021) Correlation of NDVI with RGB data to evaluate the effects of solar exposure on different combinations of ornamental grass used in lawns. In: Industrial IoT Technologies and Applications: 4th EAI International Conference, Industrial IoT 2020, Virtual Event, Proceedings 4, Springer International Publishing, 207–220. https://doi.org/10.1007/978-3-030-71061-3_13 |
| [50] |
Sishodia RP, Ray RL, Singh SK (2020) Applications of remote sensing in precision agriculture: A review. Remote Sens 12: 1–31. https://doi.org/10.3390/rs12193136 doi: 10.3390/rs12193136
|
| [51] |
Elazab A, Ordóñez RA, Savin R, et al. (2016) Detecting interactive effects of N fertilization and heat stress on maize productivity by remote sensing techniques. Eur J Agron 73: 11–24. https://doi.org/10.1016/j.eja.2015.11.010 doi: 10.1016/j.eja.2015.11.010
|
| [52] |
Bajwa SG, Rupe JC, Mason J (2017) Soybean disease monitoring with leaf reflectance. Remote Sens 9: 127. https://doi.org/10.3390/rs9020127 doi: 10.3390/rs9020127
|
| [53] |
Bhat SA, Huang NF (2021) Big data and AI revolution in precision agriculture: Survey and challenges. IEEE Access 9: 110209–110222. https://doi.org/10.1109/ACCESS.2021.3102227 doi: 10.1109/ACCESS.2021.3102227
|
| [54] |
Gutiérrez F, Htun NN, Schlenz F, et al. (2019) A review of visualisations in agricultural decision support systems: An HCI perspective. Comput Electron Agric 163: 104844. https://doi.org/10.1016/j.compag.2019.05.053 doi: 10.1016/j.compag.2019.05.053
|
| [55] |
Durak G, Ozkeskin EE, Ataizi M (2016) QR codes in education and communication. Turk Online J Distance Educ 17: 43–58. https://doi.org/10.17718/tojde.89156 doi: 10.17718/tojde.89156
|




Figures(5)

Ayomikun D. Ajayi, Boris Boiarskii, Kouya Aoyagi, Hideo Hasegawa. Utilizing MapBox API, Java, and ICT in the creation of agricultural interactive maps for improved farm management and decision-making[J]. AIMS Agriculture and Food, 2024, 9(2): 393-410. doi: 10.3934/agrfood.2024023







 DownLoad:
DownLoad: