Citation: Sergey Y. Morozov, Andrey G. Solovyev. Small hydrophobic viral proteins involved in intercellular movement of diverse plant virus genomes[J]. AIMS Microbiology, 2020, 6(3): 305-329. doi: 10.3934/microbiol.2020019

| [1] |
Yadav SR, Yan D, Sevilem I, et al. (2014) Plasmodesmata-mediated intercellular signaling during plant growth and development. Front Plant Sci 5: 44. doi: 10.3389/fpls.2014.00044

|
| [2] |
Knox K, Wang P, Kriechbaumer V, et al. (2015) Putting the squeeze on plasmodesmata: a role for reticulons in primary Plasmodesmata formation. Plant Physiol 168: 1563-1572. doi: 10.1104/pp.15.00668
|
| [3] |
Tilsner J, Nicolas W, Rosado A, et al. (2016) Staying tight: plasmodesmal membrane contact sites and the control of cell-to-cell connectivity in plants. Annu Rev Plant Biol 67: 337-364. doi: 10.1146/annurev-arplant-043015-111840
|
| [4] |
Han X, Huang LJ, Feng D, et al. (2019) Plasmodesmata-related structural and functional proteins: the long sought-after secrets of a cytoplasmic channel in plant cell walls. Int J Mol Sci 20: 2946. doi: 10.3390/ijms20122946
|
| [5] |
Dorokhov YL, Ershova NM, Sheshukova EV, et al. (2019) Plasmodesmata conductivity regulation: a mechanistic model. Plants (Basel) 8: 595. doi: 10.3390/plants8120595
|
| [6] |
Pankratenko AV, Atabekova AK, Morozov SY, et al. (2020) Membrane contacts in plasmodesmata: structural components and their functions. Biochemistry (Moscow) 85: 531-544. doi: 10.1134/S0006297920050028
|
| [7] |
Levy A, Tilsner J (2020) Creating contacts between replication and movement at plasmodesmata–a role for membrane contact sites in plant virus infections? Front Plant Sci 11: 862. doi: 10.3389/fpls.2020.00862
|
| [8] |
Sager RE, Lee JY (2018) Plasmodesmata at a glance. J Cell Sci 131: jcs209346. doi: 10.1242/jcs.209346
|
| [9] |
Pitzalis N, Heinlein M (2017) The roles of membranes and associated cytoskeleton in plant virus replication and cell-to-cell movement. J Exp Bot 69: 117-132. doi: 10.1093/jxb/erx334
|
| [10] |
Petit JD, Li ZP, Nicolas WJ, et al. (2020) Dare to change, the dynamics behind plasmodesmata-mediated cell-to-cell communication. Curr Opin Plant Biol 53: 80-89. doi: 10.1016/j.pbi.2019.10.009
|
| [11] |
Reagan BC, Burch-Smith TM (2020) Viruses reveal the secrets of plasmodesmal cell biology. Mol Plant-Microbe Interact 33: 26-39. doi: 10.1094/MPMI-07-19-0212-FI
|
| [12] |
Heinlein M (2015) Plant virus replication and movement. Virology 480: 657-671. doi: 10.1016/j.virol.2015.01.025
|
| [13] |
Lee J-Y (2015) Plasmodesmata: A signaling hub at the cellular boundary. Curr Opin Plant Biol 27: 133-140. doi: 10.1016/j.pbi.2015.06.019
|
| [14] |
Peiró A, Martínez-Gil L, Tamborero S, et al. (2014) The Tobacco mosaic virus movement protein associates with but does not integrate into biological membranes. J Virol 88: 3016-3026. doi: 10.1128/JVI.03648-13
|
| [15] |
Ishikawa K, Tamura K, Fukao Y, et al. (2020) Structural and functional relationships between plasmodesmata and plant endoplasmic reticulum-plasma membrane contact sites consisting of three synaptotagmins. New Phytol 226: 798-808. doi: 10.1111/nph.16391
|
| [16] |
Kitagawa M, Jackson D (2017) Plasmodesmata-mediated cell-to-cell communication in the shoot apical meristem: how stem cells talk. Plants (Basel) 6: 12. doi: 10.3390/plants6010012
|
| [17] | Reagan BC, Ganusova EE, Fernandez JC, et al. (2018) RNA on the move: The plasmodesmata perspective. Plant Sci Int J Exp Plant Biol 275: 1-10. |
| [18] | Navarro JA, Sanchez-Navarro JA, Pallas V (2019) Key checkpoints in the movement of plant viruses through the host. Advances in Virus Research London: Academic Press, 1-64. |
| [19] |
Atabekov JG, Morozov SY (1979) Translation of plant virus messenger RNAs. Adv Virus Res 25: 1-91. doi: 10.1016/S0065-3527(08)60568-0
|
| [20] |
Atabekov JG, Dorokhov YL (1984) Plant virus-specific transport function and resistance of plants to viruses. Adv Virus Res 29: 313-364. doi: 10.1016/S0065-3527(08)60412-1
|
| [21] |
Atabekov JG, Taliansky ME (1990) Expression of a plant virus-coded transport function by different viral genomes. Adv Virus Res 38: 201-248. doi: 10.1016/S0065-3527(08)60863-5
|
| [22] |
Deom CM, Lapidot M, Beachy RN (1992) Plant virus movement proteins. Cell 69: 221-224. doi: 10.1016/0092-8674(92)90403-Y
|
| [23] |
Leonard DA, Zaitlin M (1982) A temperature-sensitive strain of tobacco mosaic virus defective in cell-to-cell movement generates an altered viral-coded protein. Virology 117: 416-424. doi: 10.1016/0042-6822(82)90480-9
|
| [24] |
Takamatsu N, Ohno T, Meshi T, et al. (1983) Molecular cloning and nucleotide sequence of the 30K and the coat protein cistron of TMV (tomato strain) genome. Nucleic Acids Res 11: 3767-3778. doi: 10.1093/nar/11.11.3767
|
| [25] |
Meshi T, Watanabe Y, Saito T, et al. (1987) Function of the 30 kd protein of tobacco mosaic virus: involvement in cell-to-cell movement and dispensability for replication. EMBO J 6: 2557-2563. doi: 10.1002/j.1460-2075.1987.tb02544.x
|
| [26] |
Deom CM, Oliver MJ, Beachy RN (1987) The 30-kilodalton gene product of tobacco mosaic virus potentiates virus movement. Science 237: 389-394. doi: 10.1126/science.237.4813.389
|
| [27] |
Kumar D, Kumar R, Hyun TK, et al. (2015) Cell-to-cell movement of viruses via plasmodesmata. J Plant Res 128: 37-47. doi: 10.1007/s10265-014-0683-6
|
| [28] |
Ueki S, Citovsky V (2014) Plasmodesmata-associated proteins: can we see the whole elephant? Plant Signal Behav 9: e27899. doi: 10.4161/psb.27899
|
| [29] |
Yuan C, Lazarowitz SG, Citovsky V (2018) The plasmodesmal localization signal of TMV MP is recognized by plant synaptotagmin SYTA. mBio 9: e01314-e01318. doi: 10.1128/mBio.01314-18
|
| [30] |
Mushegian AR, Elena SF (2015) Evolution of plant virus movement proteins from the 30K superfamily and of their homologs integrated in plant genomes. Virology 476: 304-315. doi: 10.1016/j.virol.2014.12.012
|
| [31] |
den Hollander PW, Kieper SN, Borst JW, et al. (2016) The role of plasmodesma-located proteins in tubule-guided virus transport is limited to the plasmodesmata. Arch Virol 161: 2431-2440. doi: 10.1007/s00705-016-2936-2
|
| [32] |
Belval L, Marmonier A, Schmitt-Keichinger C, et al. (2019) From a movement-deficient grapevine fanleaf virus to the identification of a new viral determinant of nematode transmission. Viruses 11: 1146. doi: 10.3390/v11121146
|
| [33] |
Schoelz JE, Leisner S (2017) Setting up shop: The formation and function of the viral factories of cauliflower mosaic virus. Front Plant Sci 8: 1832. doi: 10.3389/fpls.2017.01832
|
| [34] |
Rodriguez A, Angel CA, Lutz L, et al. (2014) Association of the P6 protein of Cauliflower mosaic virus with plasmodesmata and plasmodesmal proteins. Plant Physiol 166: 1345-1358. doi: 10.1104/pp.114.249250
|
| [35] |
Schoelz JE, Angel CA, Nelson RS, et al. (2016) A model for intracellular movement of Cauliflower mosaic virus: the concept of the mobile virion factory. J Exp Bot 67: 2039-2048. doi: 10.1093/jxb/erv520
|
| [36] |
Hohn T (2013) Plant pararetroviruses: interactions of cauliflower mosaic virus with plants and insects. Curr Opin Virol 3: 629-638. doi: 10.1016/j.coviro.2013.08.014
|
| [37] |
Geldreich A, Haas G, Kubina J, et al. (2017) Formation of large viroplasms and virulence of Cauliflower mosaic virus in turnip plants depend on the N-terminal EKI sequence of viral protein TAV. PLoS One 12: e0189062. doi: 10.1371/journal.pone.0189062
|
| [38] |
Li JT, Yeh YC, Yeh SD, et al. (2011) Complete genomic sequence of watermelon bud necrosis virus. Arch Virol 156: 359-362. doi: 10.1007/s00705-010-0881-z
|
| [39] |
Borniego MB, Karlin D, Peña EJ, et al. (2016) Bioinformatic and mutational analysis of ophiovirus movement proteins, belonging to the 30K superfamily. Virology 498: 172-180. doi: 10.1016/j.virol.2016.08.027
|
| [40] |
Margaria P, Anderson CT, Turina M, et al. (2016) Identification of Ourmiavirus 30K movement protein amino acid residues involved in symptomatology, viral movement, subcellular localization and tubule formation. Mol Plant Pathol 17: 1063-1079. doi: 10.1111/mpp.12348
|
| [41] |
Robles Luna G, Peña EJ, Borniego MB, et al. (2018) Citrus psorosis virus movement protein contains an aspartic protease required for autocleavage and the formation of tubule-like structures at plasmodesmata. J Virol 92: e00355-18. doi: 10.1128/JVI.00355-18
|
| [42] |
Grangeon R, Jiang J, Wan J, et al. (2013) 6K2-induced vesicles can move cell to cell during turnip mosaic virus infection. Front Microbiol 4: 351. doi: 10.3389/fmicb.2013.00351
|
| [43] |
Revers F, García JA (2015) Molecular biology of potyviruses. Adv Virus Res 92: 101-109. doi: 10.1016/bs.aivir.2014.11.006
|
| [44] |
Movahed N, Patarroyo C, Sun J, et al. (2017) Cylindrical inclusion protein of turnip mosaic virus serves as a docking point for the intercellular movement of viral replication vesicles. Plant Physiol 175: 1732-1744. doi: 10.1104/pp.17.01484
|
| [45] |
Bak A, Folimonova SY (2015) The conundrum of a unique protein encoded by citrus tristeza virus that is dispensable for infection of most hosts yet shows characteristics of a viral movement protein. Virology 485: 86-95. doi: 10.1016/j.virol.2015.07.005
|
| [46] |
Qiao W, Medina V, Kuo YW, et al. (2018) A distinct, non-virion plant virus movement protein encoded by a crinivirus essential for systemic infection. mBio 9: e02230-18. doi: 10.1128/mBio.02230-18
|
| [47] |
Dolja VV, Kreuze JF, Valkonen JP (2006) Comparative and functional genomics of closteroviruses. Virus Res 117: 38-51. doi: 10.1016/j.virusres.2006.02.002
|
| [48] |
Dolja VV, Koonin EV (2013) The closterovirus-derived gene expression and RNA interference vectors as tools for research and plant biotechnology. Front Microbiol 4: 83. doi: 10.3389/fmicb.2013.00083
|
| [49] |
Morozov SY, Lukasheva LI, Chernov BK, et al. (1987) Nucleotide sequence of the open reading frames adjacent to the coat protein cistron in potato virus X genome. FEBS Lett 213: 438-442. doi: 10.1016/0014-5793(87)81538-7
|
| [50] |
Morozov S, Dolja VV, Atabekov JG (1989) Probable reassortment of genomic elements among elongated RNA-containing plant viruses. J Mol Evol 29: 52-62. doi: 10.1007/BF02106181
|
| [51] | Hull R (2002) Plant to plant movement. Plant Virology London: Academic Press, 669-751. |
| [52] |
Morozov SY, Solovyev AG (2003) Triple gene block: modular design of a multi-functional machine for plant virus movement. J Gen Virol 84: 1351-1366. doi: 10.1099/vir.0.18922-0
|
| [53] |
Genoves A, Navarro JA, Pallas V (2006) Functional analysis of the five melon necrotic spot virus genome-encoded proteins. J Gen Virol 87: 2371-2380. doi: 10.1099/vir.0.81793-0
|
| [54] |
Verchot-Lubicz J, Torrance L, Solovyev AG, et al. (2010) Varied movement strategies employed by triple gene block-encoding viruses. Mol Plant Microbe Interact 23: 1231-1247. doi: 10.1094/MPMI-04-10-0086
|
| [55] |
Solovyev AG, Kalinina NO, Morozov SY (2012) Recent advances in research of plant virus movement mediated by triple gene block. Front Plant Sci 3: 276. doi: 10.3389/fpls.2012.00276
|
| [56] |
Liou MR, Hu CC, Chou YL, et al. (2015) Viral elements and host cellular proteins in intercellular movement of Bamboo mosaic virus. Curr Opin Virol 12: 99-108. doi: 10.1016/j.coviro.2015.04.005
|
| [57] |
Navarro JA, Pallás V (2017) An update on the intracellular and intercellular trafficking of carmoviruses. Front Plant Sci 8: 1801. doi: 10.3389/fpls.2017.01801
|
| [58] |
Lazareva EA, Lezzhov AA, Komarova TV, et al. (2017) A novel block of plant virus movement genes. Mol Plant Pathol 18: 611-624. doi: 10.1111/mpp.12418
|
| [59] |
Mäkinen K, Hafrén A (2014) Intracellular coordination of potyviral RNA functions in infection. Front Plant Sci 5: 110. doi: 10.3389/fpls.2014.00110
|
| [60] |
Ivanov KI, Eskelin K, Lõhmus A, et al. (2014) Molecular and cellular mechanisms underlying potyvirus infection. J Gen Virol 95: 1415-1429. doi: 10.1099/vir.0.064220-0
|
| [61] |
Olspert A, Carr JP, Firth AE (2016) Mutational analysis of the Potyviridae transcriptional slippage site utilized for expression of the P3N-PIPO and P1N-PISPO proteins. Nucleic Acids Res 44: 7618-7629. doi: 10.1093/nar/gkw441
|
| [62] |
Valli AA, Gallo A, Rodamilans B, et al. (2018) The HCPro from the Potyviridae family: An enviable multitasking Helper Component that every virus would like to have. Mol Plant Pathol 19: 744-763. doi: 10.1111/mpp.12553
|
| [63] |
Shen WT, Wang MQ, Yan P, et al. (2010) Protein interaction matrix of Papaya ringspot virus type P based on a yeast two-hybrid system. Acta Virol 54: 49-54. doi: 10.4149/av_2010_01_49
|
| [64] |
Gibbs AJ, Hajizadeh M, Ohshima K, et al. (2020) The potyviruses: an evolutionary synthesis is emerging. Viruses 12: 132. doi: 10.3390/v12020132
|
| [65] |
Shen W, Shi Y, Dai Z, et al. (2020) The RNA-dependent rna polymerase nib of potyviruses plays multifunctional, contrasting roles during viral infection. Viruses 12: 77. doi: 10.3390/v12010077
|
| [66] |
den Boon JA, Ahlquist P (2010) Organelle-like membrane compartmentalization of positive-strand RNA virus replication factories. Annu Rev Microbiol 64: 241-256. doi: 10.1146/annurev.micro.112408.134012
|
| [67] |
Laliberté JF, Zheng H (2014) Viral manipulation of plant host membranes. Annu Rev Virol 1: 237-259. doi: 10.1146/annurev-virology-031413-085532
|
| [68] |
Paul D, Bartenschlager R (2013) Architecture and biogenesis of plus-strand RNA virus replication factories. World J Virol 2: 32-48. doi: 10.5501/wjv.v2.i2.32
|
| [69] |
Gushchin VA, Solovyev AG, Erokhina TN, et al. (2013) Beet yellows virus replicase and replicative compartments: parallels with other RNA viruses. Front Microbiol 4: 38. doi: 10.3389/fmicb.2013.00038
|
| [70] |
Jin X, Cao X, Wang X, et al. (2018) Three-dimensional architecture and biogenesis of membrane structures associated with plant virus replication. Front Plant Sci 9: 57. doi: 10.3389/fpls.2018.00057
|
| [71] | Wolff G, Melia CE, Snijder EJ, et al. (2020) Double-membrane vesicles as platforms for viral replication. Trends Microbiol 11. |
| [72] |
Chai M, Wu X, Liu J, et al. (2020) P3N-PIPO interacts with P3 via the shared N-Terminal domain to recruit viral replication vesicles for cell-to-cell movement. J Virol 94: e01898-19. doi: 10.1128/JVI.01898-19
|
| [73] |
Daròs JA, Schaad MC, Carrington JC (1999) Functional analysis of the interaction between VPg-proteinase (NIa) and RNA polymerase (NIb) of tobacco etch potyvirus, using conditional and suppressor mutants. J Virol 73: 8732-8740. doi: 10.1128/JVI.73.10.8732-8740.1999
|
| [74] |
Wei T, Wang A (2008) Biogenesis of cytoplasmic membranous vesicles for plant potyvirus replication occurs at endoplasmic reticulum exit sites in a COPI- and COPII-dependent manner. J Virol 82: 12252-12264. doi: 10.1128/JVI.01329-08
|
| [75] |
Jiang J, Patarroyo C, Garcia Cabanillas D, et al. (2015) The vesicle-forming 6K2 protein of turnip mosaic virus interacts with the COPII coatomer Sec24a for viral systemic infection. J Virol 89: 6695-6710. doi: 10.1128/JVI.00503-15
|
| [76] |
Movahed N, Sun J, Vali H, et al. (2019) A host ER fusogen is recruited by Turnip Mosaic Virus for maturation of viral replication vesicles. Plant Physiol 179: 507-518. doi: 10.1104/pp.18.01342
|
| [77] |
Cui H, Wang A (2016) Plum pox virus 6K1 protein is required for viral replication and targets the viral replication complex at the early stage of infection. J Virol 90: 5119-5131. doi: 10.1128/JVI.00024-16
|
| [78] |
Geng C, Yan ZY, Cheng DJ, et al. (2017) Tobacco vein banding mosaic virus 6K2 Protein Hijacks NbPsbO1 for Virus Replication. Sci Rep 7: 43455. doi: 10.1038/srep43455
|
| [79] |
Wei T, Zhang C, Hong J, et al. (2010) Formation of complexes at plasmodesmata for potyvirus intercellular movement is mediated by the viral protein P3N-PIPO. PLoS Pathog 6: e1000962. doi: 10.1371/journal.ppat.1000962
|
| [80] |
Wan J, Cabanillas DG, Zheng H, et al. (2015) Turnip mosaic virus moves systemically through both phloem and xylem as membrane-associated complexes. Plant Physiol 167: 1374-1388. doi: 10.1104/pp.15.00097
|
| [81] | Wu G, Cui X, Chen H, et al. (2018) Dynamin-like proteins of endocytosis in plants are coopted by potyviruses to enhance virus infection. J Virol 92: e01320-18. |
| [82] |
Lerich A, Langhans M, Sturm S, et al. (2011) Is the 6 kDa tobacco etch viral protein a bona fide ERES marker? J Exp Bot 62: 5013-5023. doi: 10.1093/jxb/err200
|
| [83] |
Cui X, Lu L, Wang Y, et al. (2018) The interaction of soybean reticulon homology domain protein (GmRHP) with Soybean mosaic virus encoded P3 contributes to the viral infection. Biochem Biophys Res Commun 495: 2105-2110. doi: 10.1016/j.bbrc.2017.12.043
|
| [84] |
Cabanillas DG, Jiang J, Movahed N, et al. (2018) Turnip nosaic virus uses the SNARE protein VTI11 in an unconventional route for replication vesicle trafficking. Plant Cell 30: 2594-2615. doi: 10.1105/tpc.18.00281
|
| [85] |
Cheng G, Dong M, Xu Q, et al. (2017) Dissecting the molecular mechanism of the subcellular localization and cell-to-cell movement of the sugarcane mosaic virus P3N-PIPO. Sci Rep 7: 9868. doi: 10.1038/s41598-017-10497-6
|
| [86] |
Sorel M, Garcia JA, German-Retana S (2014) The Potyviridae cylindrical inclusion helicase: a key multipartner and multifunctional protein. Mol Plant Microbe Interact 27: 215-226. doi: 10.1094/MPMI-11-13-0333-CR
|
| [87] |
Chen T, Ji D, Tian S (2018) Variable-angle epifluorescence microscopy characterizes protein dynamics in the vicinity of plasma membrane in plant cells. BMC Plant Biol 18: 43. doi: 10.1186/s12870-018-1246-0
|
| [88] |
Huang D, Sun Y, Ma Z, et al. (2019) Salicylic acid-mediated plasmodesmal closure via Remorin-dependent lipid organization. Proc Natl Acad Sci USA 116: 21274-21284. doi: 10.1073/pnas.1911892116
|
| [89] |
Jaillais Y, Ott T (2020) The nanoscale organization of the plasma membrane and its importance in signaling: a proteolipid perspective. Plant Physiol 182: 1682-1696. doi: 10.1104/pp.19.01349
|
| [90] |
Cheng G, Yang Z, Zhang H, et al. (2020) Remorin interacting with PCaP1 impairs Turnip mosaic virus intercellular movement but is antagonised by VPg. New Phytol 225: 2122-2139. doi: 10.1111/nph.16285
|
| [91] |
Spetz C, Valkonen JP (2004) Potyviral 6K2 protein long-distance movement and symptom-induction functions are independent and host-specific. Mol Plant Microbe Interact 17: 502-510. doi: 10.1094/MPMI.2004.17.5.502
|
| [92] |
Wan J, Laliberté JF (2015) Membrane-associated virus replication complexes locate to plant conducting tubes. Plant Signal Behav 10: e1042639. doi: 10.1080/15592324.2015.1042639
|
| [93] |
González R, Wu B, Li X, et al. (2019) Mutagenesis scanning uncovers evolutionary constraints on tobacco etch potyvirus membrane-associated 6K2 protein. Genome Biol Evol 11: 1207-1222. doi: 10.1093/gbe/evz069
|
| [94] |
Zhang L, Shang J, Jia Q, et al. (2019) The complete genome sequence of wild tomato mosaic virus isolated from Solanum nigrum reveals recombination in the P1 cistron. Arch Virol 164: 903-906. doi: 10.1007/s00705-019-04146-8
|
| [95] |
Lõhmus A, Varjosalo M, Mäkinen K (2016) Protein composition of 6K2-induced membrane structures formed during Potato virus A infection. Mol Plant Pathol 17: 943-958. doi: 10.1111/mpp.12341
|
| [96] |
Mueller BK, Subramaniam S, Senes A (2014) A frequent, GxxxG-mediated, transmembrane association motif is optimized for the formation of interhelical Cα-H hydrogen bonds. Proc Natl Acad Sci USA 111: E888-895. doi: 10.1073/pnas.1319944111
|
| [97] |
Teese MG, Langosch D (2015) Role of GxxxG motifs in transmembrane domain interactions. Biochemistry 54: 5125-5135. doi: 10.1021/acs.biochem.5b00495
|
| [98] |
Anderson SM, Mueller BK, Lange EJ, et al. (2017) Combination of Cα-H hydrogen bonds and Van der Waals packing modulates the stability of gxxxg-mediated dimers in membranes. J Am Chem Soc 139: 15774-15783. doi: 10.1021/jacs.7b07505
|
| [99] |
Fink A., Sal-Man N., Gerber D, et al. (2012) Transmembrane domains interactions within the membrane milieu: principles, advances and challenges. Biochim Biophys Acta 1818: 974-983. doi: 10.1016/j.bbamem.2011.11.029
|
| [100] |
Hu J, Rapoport TA (2016) Fusion of the endoplasmic reticulum by membrane-bound GTPases. Semin Cell Dev Biol 60: 105-111. doi: 10.1016/j.semcdb.2016.06.001
|
| [101] |
Dolja VV, Kreuze JF, Valkonen JP (2006) Comparative and functional genomics of closteroviruses. Virus Res 117: 38-51. doi: 10.1016/j.virusres.2006.02.002
|
| [102] | Kiss ZA, Medina V, Falk BW (2013) Crinivirus replication and host interactions. Front Microbiol 4: 99. |
| [103] |
Agranovsky AA (2016) Closteroviruses: molecular biology, evolution and interactions with cells. Plant viruses: evolution and management Singapore: Springer, 231-252. doi: 10.1007/978-981-10-1406-2_14
|
| [104] |
Folimonova SY (2020) Citrus tristeza virus: A large RNA virus with complex biology turned into a valuable tool for crop protection. PLoS Pathog 16: e1008416. doi: 10.1371/journal.ppat.1008416
|
| [105] | Candresse T, Fuchs M (2020) Closteroviridae. eLS . |
| [106] |
Saberi A, Gulyaeva AA, Brubacher JL, et al. (2018) A planarian nidovirus expands the limits of RNA genome size. PLoS Pathog 14: e1007314. doi: 10.1371/journal.ppat.1007314
|
| [107] |
Dolja VV, Haldeman R, Robertson NL, et al. (1994) Distinct functions of capsid protein in assembly and movement of tobacco etch potyvirus in plants. EMBO J 13: 1482-1491. doi: 10.1002/j.1460-2075.1994.tb06403.x
|
| [108] |
Kreuze JF, Savenkov EI, Valkonen JP (2002) Complete genome sequence and analyses of the subgenomic RNAs of sweet potato chlorotic stunt virus reveal several new features for the genus Crinivirus. J Virol 76: 9260-9270. doi: 10.1128/JVI.76.18.9260-9270.2002
|
| [109] |
Ling KS, Zhu HY, Gonsalves D (2004) Complete nucleotide sequence and genome organization of Grapevine leafroll-associated virus 3, type member of the genus Ampelovirus. J Gen Virol 85: 2099-2102. doi: 10.1099/vir.0.80007-0
|
| [110] |
Dolja VV, Koonin EV (2013) The closterovirus-derived gene expression and RNA interference vectors as tools for research and plant biotechnology. Front Microbiol 4: 83. doi: 10.3389/fmicb.2013.00083
|
| [111] |
Dolja VV (2003) Beet yellows virus: the importance of being different. Mol Plant Pathol 4: 91-98. doi: 10.1046/j.1364-3703.2003.00154.x
|
| [112] |
Solovyev AG, Makarov VV (2016) Helical capsids of plant viruses: architecture with structural lability. J Gen Virol 97: 1739-1754. doi: 10.1099/jgv.0.000524
|
| [113] |
Medina V, Peremyslov VV, Hagiwara Y, et al. (1999) Subcellular localization of the HSP70-homolog encoded by beet yellows closterovirus. Virology 260: 173-181. doi: 10.1006/viro.1999.9807
|
| [114] |
Peremyslov VV, Pan YW, Dolja VV (2004) Movement protein of a closterovirus is a type III integral transmembrane protein localized to the endoplasmic reticulum. J Virol 78: 3704-3709. doi: 10.1128/JVI.78.7.3704-3709.2004
|
| [115] |
Qiao W, Medina V, Falk BW (2017) Inspirations on virus replication and cell-to-cell movement from studies examining the cytopathology induced by lettuce infectious yellows virus in plant cells. Front Plant Sci 8: 1672. doi: 10.3389/fpls.2017.01672
|
| [116] |
Qiao W, Helpio EL, Falk BW (2018) Two crinivirus-conserved small proteins, P5 and P9, are indispensable for efficient lettuce infectious yellows virus infectivity in plants. Viruses 10: 459. doi: 10.3390/v10090459
|
| [117] |
Bak A, Folimonova SY (2015) The conundrum of a unique protein encoded by citrus tristeza virus that is dispensable for infection of most hosts yet shows characteristics of a viral movement protein. Virology 485: 86-95. doi: 10.1016/j.virol.2015.07.005
|
| [118] |
Wei Y, Shi Y, Han X, et al. (2019) Identification of cucurbit chlorotic yellows virus P4.9 as a possible movement protein. Virol J 16: 82. doi: 10.1186/s12985-019-1192-y
|
| [119] |
Livieratos IC, Eliasco E, Müller G, et al. (2004) Analysis of the RNA of Potato yellow vein virus: evidence for a tripartite genome and conserved 3′-terminal structures among members of the genus Crinivirus. J Gen Virol 85: 2065-2075. doi: 10.1099/vir.0.79910-0
|
| [120] |
Ruiz L, Simón A, García C, et al. (2018) First natural crossover recombination between two distinct species of the family Closteroviridae leads to the emergence of a new disease. PLoS One 13: e0198228. doi: 10.1371/journal.pone.0198228
|
| [121] |
Hadad L, Luria N, Smith E, et al. (2019) Lettuce chlorosis virus disease: a new threat to cannabis production. Viruses 11: 802. doi: 10.3390/v11090802
|
| [122] |
Martelli GP, Adams MJ, Kreuze JF, et al. (2007) Family Flexiviridae: a case study in virion and genome plasticity. Annu Rev Phytopathol 45: 73-100. doi: 10.1146/annurev.phyto.45.062806.094401
|
| [123] | Morozov SY, Agranovsky AA (2020) Alphaflexiviruses (Alphaflexiviridae). Ref Module Life Sci . |
| [124] |
Kreuze JF, Vaira AM, Menzel W, et al. (2020) ICTV virus taxonomy profile: Alphaflexiviridae. J Gen Virol 101: 699-700. doi: 10.1099/jgv.0.001436
|
| [125] |
Atsumi G, Tomita R, Yamashita T, et al. (2015) A novel virus transmitted through pollination causes ring-spot disease on gentian (Gentiana triflora) ovaries. J Gen Virol 96: 431-439. doi: 10.1099/vir.0.071498-0
|
| [126] | Savenkov EI (2020) Pomoviruses (Virgaviridae). Ref Module Life Sci . |
| [127] | Jiang Z, Yang M, Zhang Y, et al. (2020) Hordeiviruses (Virgaviridae). Ref Module Life Sci . |
| [128] | Niehl A, Liebe S, Varrelmann M, et al. (2020) Benyviruses (Benyviridae). Ref Module Life Sci . |
| [129] |
Lezzhov AA, Gushchin VA, Lazareva EA, et al. (2015) Translation of the shallot virus X TGB3 gene depends on non-AUG initiation and leaky scanning. J Gen Virol 96: 3159-3164. doi: 10.1099/jgv.0.000248
|
| [130] |
Lozano I, Leiva AM, Jimenez J, et al. (2017) Resolution of cassava-infecting alphaflexiviruses: Molecular and biological characterization of a novel group of potexviruses lacking the TGB3 gene. Virus Res 241: 53-61. doi: 10.1016/j.virusres.2017.03.019
|
| [131] |
Solovyev AG, Morozov SY (2017) Non-replicative integral membrane proteins encoded by plant alpha-like viruses: emergence of diverse orphan orfs and movement protein genes. Front Plant Sci 8: 1820. doi: 10.3389/fpls.2017.01820
|
| [132] |
Morozov SY, Solovyev AG (2015) Phylogenetic relationship of some ‘accessory’ helicases of plant positive-stranded RNA viruses: toward understanding the evolution of triple gene block. Front Microbiol 6: 508. doi: 10.3389/fmicb.2015.00508
|
| [133] |
Tamai A, Meshi T (2001) Cell-to-cell movement of potato virus X: the role of p12 and p8 encoded by the second and third open reading frames of the triple gene block. Mol Plant Microbe Interact 14: 1158-1167. doi: 10.1094/MPMI.2001.14.10.1158
|
| [134] |
Lim HS, Vaira AM, Bae H, et al. (2010) Mutation of a chloroplast-targeting signal in Alternanthera mosaic virus TGB3 impairs cell-to-cell movement and eliminates long-distance virus movement. J Gen Virol 91: 2102-2115. doi: 10.1099/vir.0.019448-0
|
| [135] |
Morozov SY, Solovyev AG (2012) Did silencing suppression counter-defensive strategy contribute to origin and evolution of the triple gene block coding for plant virus movement proteins? Front Plant Sci 3: 136. doi: 10.3389/fpls.2012.00136
|
| [136] |
Lim HS, Bragg JN, Ganesan U, et al. (2009) Subcellular localization of the barley stripe mosaic virus triple gene block proteins. J Virol 83: 9432-9448. doi: 10.1128/JVI.00739-09
|
| [137] |
Schepetilnikov MV, Solovyev AG, Gorshkova EN, et al. (2008) Intracellular targeting of a hordeiviral membrane spanning movement protein: sequence requirements and involvement of an unconventional mechanism. J Virol 82: 1284-1293. doi: 10.1128/JVI.01164-07
|
| [138] |
Zamyatnin AA, Solovyev AG, Savenkov EI, et al. (2004) Transient coexpression of individual genes encoded by the triple gene block of potato mop-top virus reveals requirements for TGBp1 trafficking. Mol Plant Microbe Interact 17: 921-930. doi: 10.1094/MPMI.2004.17.8.921
|
| [139] |
Ju HJ, Brown JE, Ye CM, et al. (2007) Mutations in the central domain of potato virus X TGBp2 eliminate granular vesicles and virus cell-to-cell trafficking. J Virol 81: 1899-1911. doi: 10.1128/JVI.02009-06
|
| [140] |
Bamunusinghe D, Hemenway CL, Nelson RS, et al. (2009) Analysis of potato virus X replicase and TGBp3 subcellular locations. Virology 393: 272-285. doi: 10.1016/j.virol.2009.08.002
|
| [141] |
Tilsner J, Linnik O, Louveaux M, et al. (2013) Replication and trafficking of a plant virus are coupled at the entrances of plasmodesmata. J Cell Biol 201: 981-995. doi: 10.1083/jcb.201304003
|
| [142] | Park MR, Jeong RD, Kim KH (2014) Understanding the intracellular trafficking and intercellular transport of potexviruses in their host plants. Front Plant Sci 5: a60. |
| [143] | Wu X, Liu J, Chai M, et al. (2019) The Potato virus X TGBp2 protein plays dual functional roles in viral replication and movement. J Virol 93: e01635-18. |
| [144] |
Chou YL, Hung YJ, Tseng YH, et al. (2013) The stable association of virion with the triple-gene-block protein 3-based complex of Bamboo mosaic virus. PLoS Pathog 9: e1003405. doi: 10.1371/journal.ppat.1003405
|
| [145] |
Linnik O, Liesche J, Tilsner J, et al. (2013) Unraveling the structure of viral replication complexes at super-resolution. Front Plant Sci 4: 6. doi: 10.3389/fpls.2013.00006
|
| [146] |
Boevink P, Oparka KJ (2005) Virus-host interactions during movement processes. Plant Physiol 138: 1815-1821. doi: 10.1104/pp.105.066761
|
| [147] |
Karpova OV, Zayakina OV, Arkhipenko MV, et al. (2006) Potato virus X RNA-mediated assembly of single-tailed ternary ‘coat protein-RNA-movement protein’ complexes. J Gen Virol 87: 2731-2740. doi: 10.1099/vir.0.81993-0
|
| [148] |
Arkhipenko MV, Nikitin NA, Donchenko EK, et al. (2017) Translational cross-activation of Potexviruses virion genomic RNAs. Acta Naturae 9: 52-57. doi: 10.32607/20758251-2017-9-4-52-57
|
| [149] |
Kalinina NO, Rakitina DV, Solovyev AG, et al. (2002) RNA helicase activity of the plant virus movement proteins encoded by the first gene of the triple gene block. Virology 296: 321-329. doi: 10.1006/viro.2001.1328
|
| [150] |
Leshchiner AD, Minina EA, Rakitina DV, et al. (2008) Oligomerization of the potato virus X 25-kD movement protein. Biochemistry (Moscow) 73: 50-55. doi: 10.1134/S0006297908010070
|
| [151] |
Howard AR, Heppler ML, Ju HJ, et al. (2004) Potato virus X TGBp1 induces plasmodesmata gating and moves between cells in several host species whereas CP moves only in N. benthamiana leaves. Virology 328: 185-197. doi: 10.1016/j.virol.2004.06.039
|
| [152] |
Lough TJ, Netzler NE, Emerson SJ, et al. (2000) Cell-to-cell movement of potexviruses: evidence for a ribonucleoprotein complex involving the coat protein and first triple gene block protein. Mol Plant Microbe Interact 13: 962-974. doi: 10.1094/MPMI.2000.13.9.962
|
| [153] |
Bayne EH, Rakitina DV, Morozov SY, et al. (2005) Cell-to-cell movement of potato potexvirus X is dependent on suppression of RNA silencing. Plant J 44: 471-482. doi: 10.1111/j.1365-313X.2005.02539.x
|
| [154] |
Voinnet O, Lederer C, Baulcombe DC (2000) A viral movement protein prevents spread of the gene silencing signal in Nicotiana benthamiana. Cell 103: 157-167. doi: 10.1016/S0092-8674(00)00095-7
|
| [155] |
Lim HS, Vaira AM, Reinsel MD, et al. (2010) Pathogenicity of Alternanthera mosaic virus is affected by determinants in RNA-dependent RNA polymerase and by reduced efficacy of silencing suppression in a movement-competent TGB1. J Gen Virol 91: 277-287. doi: 10.1099/vir.0.014977-0
|
| [156] |
Senshu H, Ozeki J, Komatsu K, et al. (2009) Variability in the level of RNA silencing suppression caused by triple gene block protein 1 (TGBp1) from various potexviruses during infection. J Gen Virol 90: 1014-1024. doi: 10.1099/vir.0.008243-0
|
| [157] |
Tilsner J, Cowan GH, Roberts AG, et al. (2010) Plasmodesmal targeting and intercellular movement of potato mop-top pomovirus is mediated by a membrane anchored tyrosine-based motif on the lumenal side of the endoplasmic reticulum and the C-terminal transmembrane domain in the TGB3 movement protein. Virology 402: 41-51. doi: 10.1016/j.virol.2010.03.008
|
| [158] |
Torrance L, Wright KM, Crutzen F, et al. (2011) Unusual features of pomoviral RNA movement. Front Microbiol 2: 259. doi: 10.3389/fmicb.2011.00259
|
| [159] |
Jackson AO, Lim HS, Bragg J, et al. (2009) Hordeivirus replication, movement, and pathogenesis. Annu Rev Phytopathol 47: 385-422. doi: 10.1146/annurev-phyto-080508-081733
|
| [160] |
Hipper C, Brault V, Ziegler-Graff V, et al. (2013) Viral and cellular factors involved in phloem transport of plant viruses. Front Plant Sci 4: a154. doi: 10.3389/fpls.2013.00154
|
| [161] |
Makarov VV, Makarova SS, Makhotenko AV, et al. (2015) In vitro properties of hordeivirus TGB1 protein forming ribonucleoprotein complexes. J Gen Virol 96: 3422-3431. doi: 10.1099/jgv.0.000252
|
| [162] |
Shemyakina EA, Solovyev AG, Leonova OG, et al. (2011) The role of microtubule association in plasmodesmal targeting of potato mop-top virus movement protein TGBp1. Open Virol J 5: 1-11. doi: 10.2174/1874357901105010001
|
| [163] |
Wright KM, Cowan GH, Lukhovitskaya NI, et al. (2010) The N-terminal domain of PMTV TGB1 movement protein is required for nucleolar localization, microtubule association, and long-distance movement. Mol Plant Microbe Interact 23: 1486-1497. doi: 10.1094/MPMI-05-10-0105
|
| [164] |
Chiba S, Hleibieh K, Delbianco A, et al. (2013) The benyvirus RNA silencing suppressor is essential for long-distance movement, requires both zinc-finger and NoLS basic residues but not a nucleolar localization for its silencing-suppression activity. Mol Plant Microbe Interact 26: 168-181. doi: 10.1094/MPMI-06-12-0142-R
|
| [165] |
Gilmer D, Ratti C (2017) ICTV Virus Taxonomy Profile: Benyviridae. J Gen Virol 98: 1571-1572. doi: 10.1099/jgv.0.000864
|
| [166] |
Randles JW, Rohde W (1990) Nicotiana velutina mosaic virus: evidence for a bipartite genome comprising 3 kb and 8 kb RNAs. J Gen Virol 71: 1019-1027. doi: 10.1099/0022-1317-71-5-1019
|
| [167] |
Melzer MJ, Sether DM, Borth WB, et al. (2012) Characterization of a virus infecting Citrus volkameriana with citrus leprosis-like symptoms. Phytopathology 102: 122-127. doi: 10.1094/PHYTO-01-11-0013
|
| [168] |
Lazareva EA, Lezzhov AA, Golyshev SA, et al. (2017) Similarities in intracellular transport of plant viral movement proteins BMB2 and TGB3. J Gen Virol 98: 2379-2391. doi: 10.1099/jgv.0.000914
|
| [169] |
Nunes MRT, Contreras-Gutierrez MA, Guzman H, et al. (2017) Genetic characterization, molecular epidemiology, and phylogenetic relationships of insect-specific viruses in the taxon Negevirus. Virology 504: 152-167. doi: 10.1016/j.virol.2017.01.022
|
| [170] |
Genovés A, Navarro J, Pallás V (2006) Functional analysis of the five melon necrotic spot virus genome-encoded proteins. J Gen Virol 87: 2371-2380. doi: 10.1099/vir.0.81793-0
|
| [171] |
Martínez-Turiño S, Hernández C (2011) A membrane-associated movement protein of Pelargonium flower break virus shows RNA-binding activity and contains a biologically relevant leucine zipper-like motif. Virology 413: 310-319. doi: 10.1016/j.virol.2011.03.001
|
| [172] |
Li WZ, Qu F, Morris TJ (1998) Cell-to-cell movement of turnip crinkle virus is controlled by two small open reading frames that function in trans. Virology 244: 405-416. doi: 10.1006/viro.1998.9125
|
| [173] | White KA (2020) Tombusvirus-Like Viruses (Tombusviridae). Ref Module Life Sci . |
| [174] |
Vilar M, Esteve V, Pallás V, et al. (2001) Structural properties of carnation mottle virus p7 movement protein and its RNA-binding domain. J Biol Chem 276: 18122-18129. doi: 10.1074/jbc.M100706200
|
| [175] |
Vilar M, Saurí A, Marcos JF, et al. (2005) Transient structural ordering of the RNA-binding domain of carnation mottle virus p7 movement protein modulates nucleic acid binding. Chembiochem 6: 1391-1396. doi: 10.1002/cbic.200400451
|
| [176] |
Navarro JA, Serra-Soriano M, Corachán-Valencia L, et al. (2020) A conserved motif in three viral movement proteins from different genera is required for host factor recruitment and cell-to-cell movement. Sci Rep 10: 4758. doi: 10.1038/s41598-020-61741-5
|
| [177] |
Genoves A, Navarro JA, Pallas V (2009) A self-interacting carmovirus movement protein plays a role in binding of viral RNA during the cell-to-cell movement and shows an actin cytoskeleton dependent location in cell periphery. Virology 395: 133-142. doi: 10.1016/j.virol.2009.08.042
|
| [178] |
Serra-Soriano M, Pallás V, Navarro JA (2014) A model for transport of a viral membrane protein through the early secretory pathway: minimal sequence and endoplasmic reticulum lateral mobility requirements. Plant J 77: 863-879. doi: 10.1111/tpj.12435
|
| [179] |
Martínez-Gil L, Saurí A, Vilar M, et al. (2007) Membrane insertion and topology of the p7B movement protein of Melon Necrotic Spot Virus (MNSV). Virology 367: 348-357. doi: 10.1016/j.virol.2007.06.006
|
| [180] |
Genoves A, Navarro JA, Pallas V (2010) The intra- and intercellular movement of Melon necrotic spot virus (MNSV) depends on an active secretory pathway. Mol Plant-Microbe Interact 23: 263-272. doi: 10.1094/MPMI-23-3-0263
|
| [181] |
Sauri A, Saksena S, Salgado J, et al. (2005) Double-spanning plant viral movement protein integration into the endoplasmic reticulum membrane is signal recognition particle-dependent, translocon-mediated, and concerted. J Biol Chem 280: 25907-25912. doi: 10.1074/jbc.M412476200
|
| [182] |
Genovés A, Pallas V, Navarro J (2011) Contribution of topology determinants of a viral movement protein to its membrane association, intracellular traffic and viral cell-to-cell movement. J Virol 85: 7797-7809. doi: 10.1128/JVI.02465-10
|
| [183] |
Thekke-Veetil T, Ho T, Postman JD, et al. (2018) A virus in american blackcurrant (Ribes americanum) with distinct genome features reshapes classification in the Tymovirales. Viruses 10: 406. doi: 10.3390/v10080406
|
| [184] |
Miller WA, Jackson J, Feng Y (2015) Cis- and trans-regulation of luteovirus gene expression by the 3′ end of the viral genome. Virus Res 206: 37-45. doi: 10.1016/j.virusres.2015.03.009
|
| [185] |
Ali M, Hameed S, Tahir M (2014) Luteovirus: insights into pathogenicity. Arch Virol 159: 2853-2860. doi: 10.1007/s00705-014-2172-6
|
| [186] | Domier LL (2020) Luteoviruses (Luteoviridae). Ref Module Life Sci . |
| [187] | Garcia-Ruiz H, Holste NM, LaTourrette K (2020) Poleroviruses (Luteoviridae). Ref Module Life Sci . |
| [188] |
Firth AE, Brierley I (2012) Non-canonical translation in RNA viruses. J Gen Virol 93: 1385-1409. doi: 10.1099/vir.0.042499-0
|
| [189] |
Smirnova E, Firth AE, Miller WA, et al. (2015) Discovery of a small non-AUG-initiated ORF in poleroviruses and luteoviruses that is required for long-distance movement. PLoS Pathog 11: e1004868. doi: 10.1371/journal.ppat.1004868
|
| [190] |
Zhang XY, Zhao TY, Li YY, et al. (2018) The conserved proline18 in the polerovirus p3a is important for brassica yellows virus systemic infection. Front. Microbiol 9: 613. doi: 10.3389/fmicb.2018.00613
|
| [191] |
Li S, Su X, Luo X, et al. (2020) First evidence showing that Pepper vein yellows virus P4 protein is a movement protein. BMC Microbiol 20: 72. doi: 10.1186/s12866-020-01758-y
|
| [192] |
DeBlasio SL, Xu Y, Johnson RS, et al. (2018) The interaction dynamics of two potato leafroll virus movement proteins affects their localization to the outer membranes of mitochondria and plastids. Viruses 10: 585. doi: 10.3390/v10110585
|
| [193] |
Gutierrez C (2000) Geminiviruses and the plant cell cycle. Plant Mol Biol 43: 763-772. doi: 10.1023/A:1006462028363
|
| [194] |
Gronenborn B (2004) Nanoviruses: genome organisation and protein function. Vet Microbiol 98: 103-109. doi: 10.1016/j.vetmic.2003.10.015
|
| [195] |
Krupovic M, Varsani A, Kazlauskas D, et al. (2020) Cressdnaviricota: a virus phylum unifying seven families of rep-encoding viruses with single-stranded, circular DNA genomes. J Virol 94: e00582-20. doi: 10.1128/JVI.00582-20
|
| [196] |
Zhao L, Rosario K, Breitbart M, et al. (2019) Eukaryotic circular rep-encoding single-stranded dna (cress dna) viruses: ubiquitous viruses with small genomes and a diverse host range. Adv Virus Res 103: 71-133. doi: 10.1016/bs.aivir.2018.10.001
|
| [197] | Martin DP, Monjane AL (2020) Maize Streak Virus (Geminiviridae). Ref Module Life Sci . |
| [198] | Navas-Castillo J, Fiallo-Olivé E (2020) Geminiviruses (Geminiviridae). Ref Module Life Sci . |
| [199] | Vetten HJ, Gronenborn B (2020) Nanoviruses (Nanoviridae). Ref Module Life Sci . |
| [200] |
Hanley-Bowdoin L, Bejarano ER, Robertson D, et al. (2013) Geminiviruses: masters at redirecting and reprogramming plant processes. Nat Rev Microbiol 11: 777-788. doi: 10.1038/nrmicro3117
|
| [201] | Jeske H (2009) Geminiviruses. Curr Top Microbiol Immunol 331: 185-226. |
| [202] |
Fondong VN (2013) Geminivirus protein structure and function. Mol Plant Pathol 14: 635-649. doi: 10.1111/mpp.12032
|
| [203] |
Katul L, Maiss E, Morozov SY, et al. (1997) Analysis of six DNA components of the faba bean necrotic yellows virus genome and their structural affinity to related plant virus genomes. Virology 233: 247-259. doi: 10.1006/viro.1997.8611
|
| [204] |
Wanitchakorn R, Hafner GJ, Harding RM, et al. (2000) Functional analysis of proteins encoded by banana bunchy top virus DNA-4 to -6. J Gen Virol 81: 299-306. doi: 10.1099/0022-1317-81-1-299
|
| [205] |
Grigoras I, Vetten HJ, Commandeur U, et al. (2018) Nanovirus DNA-N encodes a protein mandatory for aphid transmission. Virology 522: 281-291. doi: 10.1016/j.virol.2018.07.001
|
| [206] |
Krenz B, Schiessl I, Greiner E, et al. (2017) Analyses of pea necrotic yellow dwarf virus-encoded proteins. Virus Genes 53: 454-463. doi: 10.1007/s11262-017-1439-x
|
| [207] | Lazareva EA, Lezzhov AA, Chergintsev DA, et al. (2020) Reticulon-like properties of a plant virus-encoded movement protein. New Phytol in press. |
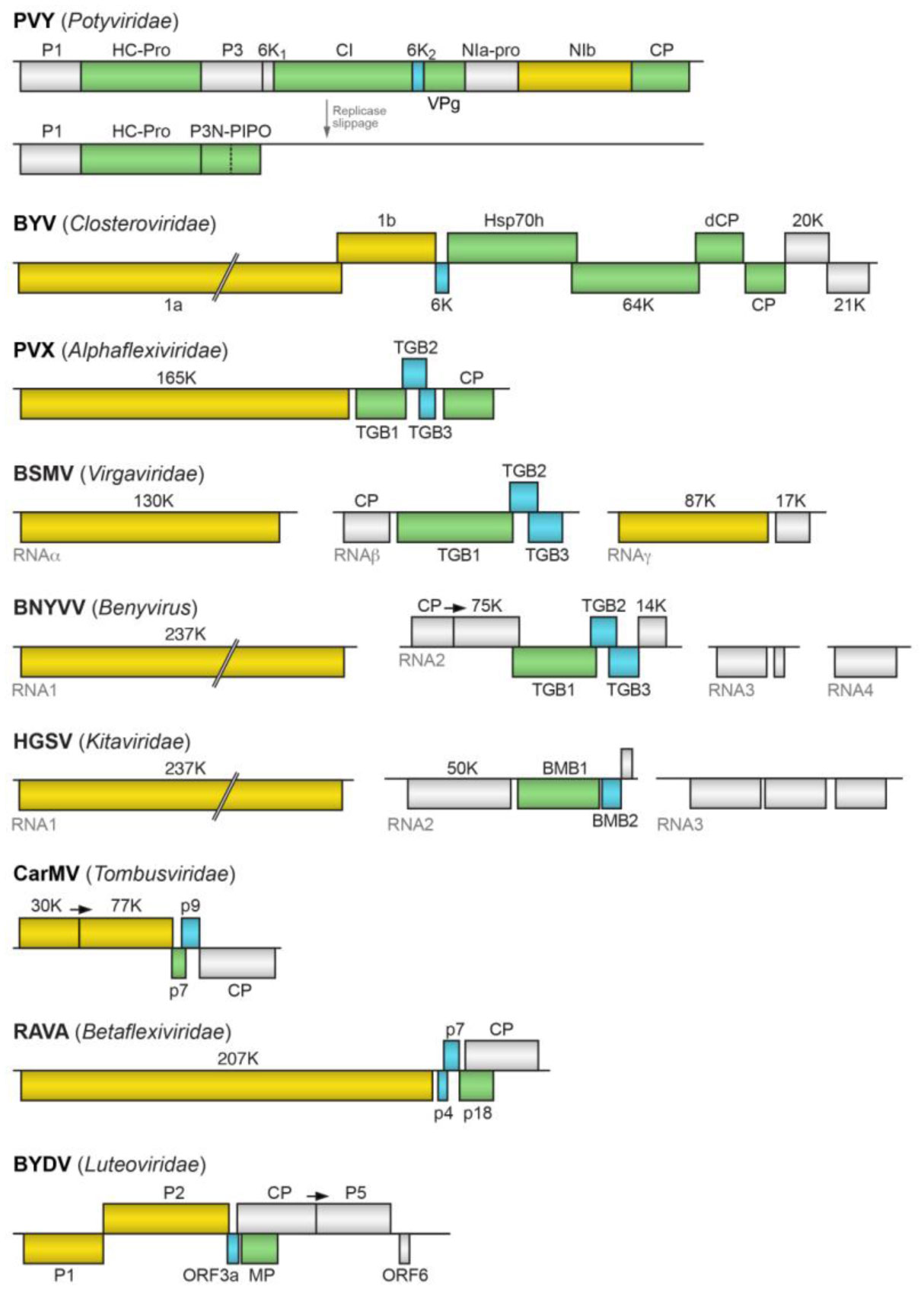
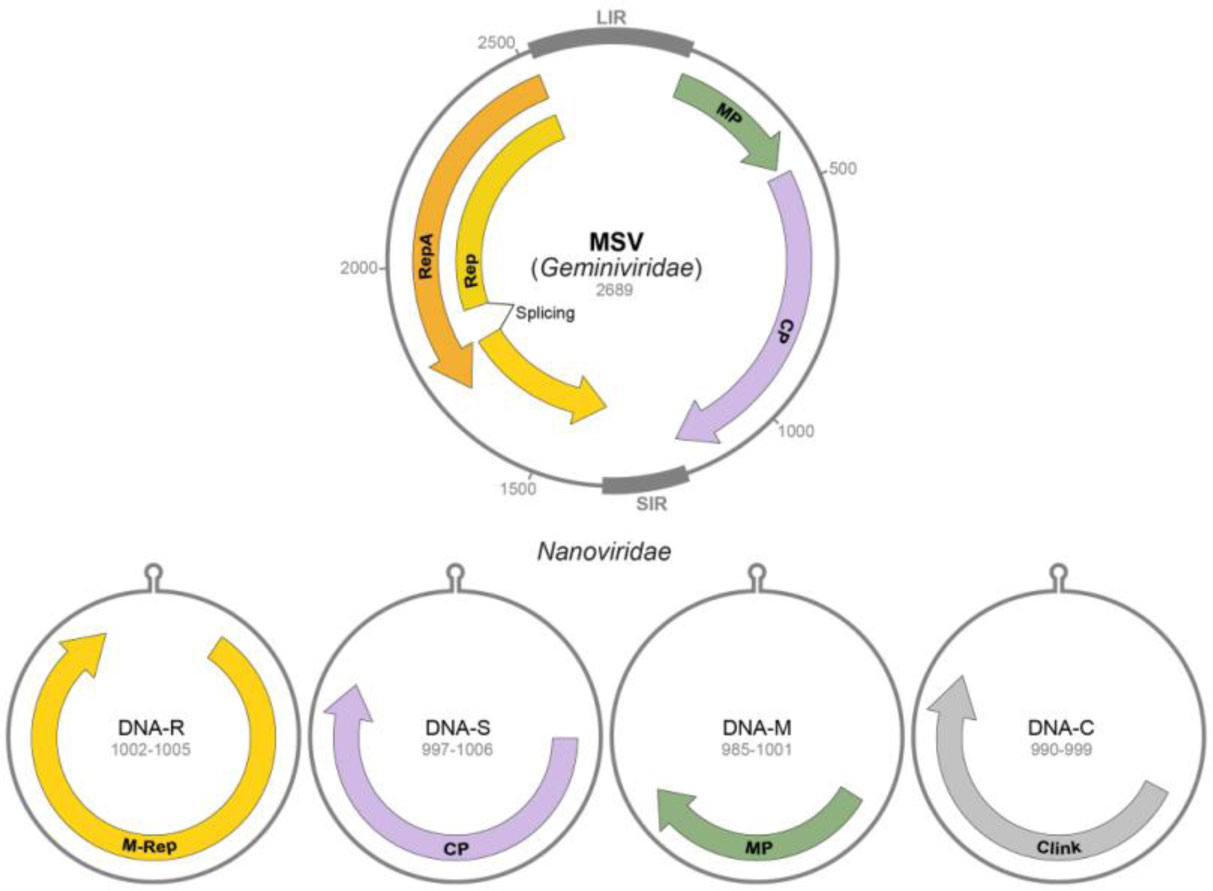
Figures(2) / Tables(1)

Sergey Y. Morozov, Andrey G. Solovyev. Small hydrophobic viral proteins involved in intercellular movement of diverse plant virus genomes[J]. AIMS Microbiology, 2020, 6(3): 305-329. doi: 10.3934/microbiol.2020019







 DownLoad:
DownLoad: