Citation: Pontsho Patricia Twala, Alfred Mitema, Cindy Baburam, Naser Aliye Feto. Breakthroughs in the discovery and use of different peroxidase isoforms of microbial origin[J]. AIMS Microbiology, 2020, 6(3): 330-349. doi: 10.3934/microbiol.2020020

| [1] |
Karigar CS, Rao SS (2011) Role of microbial enzymes in the bioremediation of pollutants: A review. Enzyme Res 2011: 805187. doi: 10.4061/2011/805187

|
| [2] | Chandrasekaran M, Kumar SR (2010) Marine microbial enzymes. Biotechnology 9: 1-15. |
| [3] | Opwis K, Kiehl K, Gutmann JS (2016) Immobilization of peroxidases on textile carrier materials and thier use in bleaching processes. Chem Eng Trans 49: 67-72. |
| [4] |
Casciello C, Tonin F, Berini F, et al. (2017) A valuable peroxidase activity from the novel species Nonomuraea gerenzanensis growing on alkali lignin. Biotechnol Rep 13: 49-57. doi: 10.1016/j.btre.2016.12.005
|
| [5] |
Madhu A, Chakraborty JN (2017) Developments in application of enzymes for textile processing. J Cleaner Prod 145: 114-133. doi: 10.1016/j.jclepro.2017.01.013
|
| [6] |
Bansal N, Kanwar SS (2013) Peroxidase(s) in environment protection. Sci World J 2013: 714639. doi: 10.1155/2013/714639
|
| [7] |
Choi J, Detry N, Kim KT, et al. (2014) fPoxDB: fungal peroxidase database for comparative genomics. BMC Microbiol 14: 117. doi: 10.1186/1471-2180-14-117
|
| [8] |
Colpa DI, Fraaije MW, Bloois EV (2014) DyP-type peroxidase: a promising versatile class of enzymes. J Ind Microbiol Biotechnol 41: 1-7. doi: 10.1007/s10295-013-1371-6
|
| [9] |
Kolhe P, Ingle S, Wagh N (2015) Degradation of phenol containing wastewater by advance catalysis system–a review. Annu Res Rev Biol 8: 1-15. doi: 10.9734/ARRB/2015/19936
|
| [10] |
Lončar N, Colpa DI, Fraaije MW (2016) Exploring the biocatalytic potential of a DyP-type peroxidase by profiling the substrate acceptance of Thermobifida fusca DyP peroxidase. Tetrahedrom 72: 7276-7281. doi: 10.1016/j.tet.2015.12.078
|
| [11] |
Pandey VP, Bhagat PK, Prajapati R, et al. (2016) A defense associated peroxidase from lemon having dye decolorizing ability and offering resistance to heat, heavy metals and organic solvents. Biochem Anal Biochem 5: 291. doi: 10.4172/2161-1009.1000291
|
| [12] |
Passardi F, Theiler G, Zamocky M, et al. (2007) PeroxiBase: the peroxidase database. Phytochemistry 68: 1605-1611. doi: 10.1016/j.phytochem.2007.04.005
|
| [13] | Rajkumar R, Yaakob Z, Takriff MS, et al. (2013) Optimization of medium composition for the production of peroxidase by Bacillus sp. Der Pharma Chemica 5: 167-174. |
| [14] |
Zhang Z, Lai J, Wu K, et al. (2018) Peroxidase-catalyzed chemiluminescence system and its application in immunoassay. Talanta 180: 260-270. doi: 10.1016/j.talanta.2017.12.024
|
| [15] | Patil SR (2014) Production and purification of lignin peroxidase from Bacillus megaterium and its application in bioremediation. CIBTech J Microbiol 3: 22-28. |
| [16] |
Adewale IO, Adekunle AT (2018) Biochemical properties of peroxidase from white and red cultivars of Kola nut (Cola nitida). Biocatal Agric Biotechnol 14: 1-9. doi: 10.1016/j.bcab.2018.01.013
|
| [17] |
Cesarino I, Araujo P, Sampaio Mayer JL, et al. (2012) Enzymatic activity and proteomic profile of class III peroxidases during sugarcane stem development. Plant Physiol Biochem 55: 66-76. doi: 10.1016/j.plaphy.2012.03.014
|
| [18] |
Li J, Liu C, Li B (2015) Identification and molecular characterization of a novel DyP-type peroxidase from Pseudomonas aeruginosa PKE117. Appl Biochem Biotechnol 166: 774-85. doi: 10.1007/s12010-011-9466-x
|
| [19] | Ganesh P, Dineshraj D, Yoganathan K (2017) Production and screening of depolymerising enymes by potential bacteria and fungi isolated from plastic waste dump yard sites. Int J Appl Res 3: 693-695. |
| [20] |
Khelil O, Choubane S, Cheba BA (2015) Co-production of cellulases and manganese peroxidases by Bacillus sp. R2 and Bacillus Cereus 11778 on waste newspaper: application in dyes decolourization. Procedia Technol 19: 980-987. doi: 10.1016/j.protcy.2015.02.140
|
| [21] |
Pandey VP, Awasthi M, Singh S, et al. (2017) A comprehensive review on function and application of plant peroxidases. Biochem Anal Biochem 6: 1. doi: 10.4172/2161-1009.1000308
|
| [22] |
Pathak VM, Navneet (2017) Review on the current status of polymer degradation: a microbial approach. Bioresources Bioprocessing 4: 15. doi: 10.1186/s40643-017-0145-9
|
| [23] |
Wei R, Zimmermann W (2017) Microbial enzymes for the recycling of recalcitrant petroleum-based plastics: how far are we? Microb Biotechnol 10: 1308-1322. doi: 10.1111/1751-7915.12710
|
| [24] |
Zahmatkesh M, Spanjers H, van Lier JB, et al. (2017) Fungal treatment of humic-rich industrial wastewater: application of white rot fungi in remediation of food-processing wastewater. Environ Technol 38: 2752-2762. doi: 10.1080/09593330.2016.1276969
|
| [25] |
Xie X, Chen M, Aiyi Z (2017) Identification and characterization of two selenium-dependent glutathione proxidase 1 isoforms from Larimichthys crocea. Fish Shellfish Immunol 71: 411-422. doi: 10.1016/j.fsi.2017.09.067
|
| [26] |
Yoshida T, Sugano Y (2015) A structural and functional perspective of DyP-type peroxidase family. Arch Biochem Biophys 574: 49-55. doi: 10.1016/j.abb.2015.01.022
|
| [27] |
Datta R, Kelkar A, Baraniya D, et al. (2017) Enzymatic degradation of lignin in soil: a review. Sustainability 9: 1163. doi: 10.3390/su9071163
|
| [28] |
Min K, Gong G, Woo H, et al. (2015) A dye-decolorizing peroxidase from Bacillus subtilis exhibiting subtrate-dependent optimum temperature for dyes and beta-ether lignin dimer. Sci Rep 5: 8245. doi: 10.1038/srep08245
|
| [29] | Bholay AD, Borkhataria BV, Jadhav PU, et al. (2012) Bacterial lignin peroxidase: a tool for biobleaching and biodegradation of industrial effluents. Universal J Environ Res Technol 2: 58-64. |
| [30] | Guisado G, Lopez MJ, Vargas-Garcia MC, et al. (2012) Pseudallescheria angusta, A lingninolytic microorganisms for wood fibres biomodification. Bioresources 7: 464-474. |
| [31] | Niladevi KN (2009) Lignolytic enzymes. Biotechnol Agro-Ind Residues Util 4: 397-414. |
| [32] |
Osuji AC, Osayi EE, Eze SOO, et al. (2014) Biobleaching of industrial important dyes with peroxidase partially purified from garlic. Scientific World J 2014: 183163. doi: 10.1155/2014/183163
|
| [33] |
Baysal O, Yildiz A (2017) Bacillus subtilis: An industrially important microbe for enzyme production. EC Microbiol 5: 148-156. doi: 10.1186/s40168-017-0368-1
|
| [34] |
Dragana R, Nikola G, Željko D, et al. (2017) Separation of peroxidases from Miscanthus x giganteus, their partial characterisation and application for degradation of dyes. Plant Physiol Biochem 120: 179-185. doi: 10.1016/j.plaphy.2017.10.009
|
| [35] |
Chanwun T, Muhamad N, Chirapongsatonkul N, et al. (2013) Hecea brasiliensis cell suspension peroxidase: purification, chracterization and application for dye decolorization. AMB Express 3: 14. doi: 10.1186/2191-0855-3-14
|
| [36] |
van Bloois E, Torres Pazmino DE, Winter RT, et al. (2010) A robust and extracellular heme-containing peroxidase from Thermobifida fusca as prototype of a bacterial peroxidase superfamily. Appl Microbiol Biotechnol 86: 1419-1430. doi: 10.1007/s00253-009-2369-x
|
| [37] |
Shigeto J, Tsutsumi Y (2016) Diverse functions and reactions of class III peroxidases. New Phytol 209: 1395-1402. doi: 10.1111/nph.13738
|
| [38] |
Veitch NC (2004) Horseradish peroxidase: a modern view of a classic enzyme. Phytochemistry 65: 249-259. doi: 10.1016/j.phytochem.2003.10.022
|
| [39] |
Petrić M, Subotić A, Jevremović S, et al. (2015) Esterase and peroxidase isoforms in different stages of morphogenesis in Fritillaria meleagris L. in bulb-scale culture. Comptes Rendus Biologies 338: 793-802. doi: 10.1016/j.crvi.2015.08.002
|
| [40] |
Racz A, Hideg E, Czegeny G (2018) Selective responses of class III plant peroxidase isoforms to environmentally relevant UV-B doses. J Plant Physiol 221: 101-106. doi: 10.1016/j.jplph.2017.12.010
|
| [41] | Kanwar SS, Bansal N (2014) Decolorization of industrial dyes by an extracellular peroxidase from Bacillus sp. F31. JAM 1: 252-265. |
| [42] | Gholami-Borujeni F, Mahvi AH, Naseri S, et al. (2011) Application of immobilized horseradish peroxidase for removal and detoxification of azo dye from aqueous solution. Res J Chem Environ 15: 217-221. |
| [43] |
Shafi J, Tian H, Ji M (2017) Bacillus species as versatile weapons for plant pathogens a review. Biotechnol Biotechnol Equipment 31: 446-459. doi: 10.1080/13102818.2017.1286950
|
| [44] |
Matamoros MA, Saiz A, Penuelas M, et al. (2015) Function of glutathion peroxidase in legume root nodules. J Exp Bot 66: 2979-2990. doi: 10.1093/jxb/erv066
|
| [45] |
Silva MC, Torres JA, Corrêa AD, et al. (2012) Obtention of plant peroxidase and its potential for the decolorization of the reactive dye Remazol Turquoise G133%. Water Sci Technol 65: 669-675. doi: 10.2166/wst.2012.892
|
| [46] | Singh R, Kumar M, Mittal A, et al. (2016) Microbial enzymes: industrial progress in 21st century. Biotechnology 6: 174. |
| [47] |
Ruiz-Dueñas F, Fernández E, Martínez MJ, et al. (2011) Pleurotus ostreatus heme peroxidases: An in silico analysis from the genome sequence to the enzyme molecular strucuture. C R Biol 334: 795-805. doi: 10.1016/j.crvi.2011.06.004
|
| [48] | Malomo SO, Adeoye RL, Babatunde L, et al. (2011) Suicide inactivation of horseradish peroxidase by excess hydrogen peroxide: the effects of reaction pH, buffer ion concentration, and redoz mediation. Biokemistri 23: 124-128. |
| [49] |
Falade A, Mabinya L, Okoh A, et al. (2019) Studies on peroxidase productin and detection of Sporotrichum thermophile-like catalase-peroxidase gene in a Bacillus species isolated from Hogsback forest reserve, South Africa. Heliyon 5: e03012. doi: 10.1016/j.heliyon.2019.e03012
|
| [50] | Simo C, Djocgoue PF, Minyaka E, et al. (2018) Guaicol proxidase heritability in tolerance of cocoa (Theobroma cacao L.) to Phytophthora megakarya, agent of cocoa black pod disease. Int J Agric Policy Res 6: 7-20. |
| [51] |
Shahin SA, Jonathan S, Lary DJ, et al. (2017) Phytophthora megakarya and Phytophthora palmivora, closely related casual agents of cacao balsk pod rot, underwent increases in genome sizes and gene numbers by differnt mechanisms. Genome Biol Evol 9: 536-557. doi: 10.1093/gbe/evx021
|
| [52] | Shahin SA, Jonathan S, Lary DJ, et al. (2017) Phytothora megakarya and P. palmivora, casual agents of black pod rot, induce similar plant defense responses late during infection of susceptible cacoa pods. Fontiers Plant Sci 18: 1-18. |
| [53] |
Falade A, Mabinya L, Okoh A, et al. (2018) Peroxidases produced by new ligninolytic Bacillus strains isolated form marsh and grassland decolourized anthraquinone and azo dyes. Pol J Environ Stud 28: 3163-3172. doi: 10.15244/pjoes/92520
|
| [54] |
Husain Q (2010) Peroxidase mediated decolorizatio and remediation of wastewar containing industrial dyes: a review. Rev Environ Sci Biotechnol 9: 117-140. doi: 10.1007/s11157-009-9184-9
|
| [55] |
Karn SK, Fang G, Duan J (2017) Bacillus sp. acting as dual role for corrosion induction and corrosion inhibition with carbon steel (CS). Front Microbiol 8: 2038. doi: 10.3389/fmicb.2017.02038
|
| [56] |
Nayanashree G, Thippeswamy B (2015) Natural rubber degradation by laccase and manganese peroxidase enzymes of Penicillium chrysogenum. Int J Environ Sci Technol 12: 2665-2672. doi: 10.1007/s13762-014-0636-6
|
| [57] | Nayanashree G, Thippeswamy B, Krishnappa M., et al. (2014) Enzymatic studies on Natural rubber biodegradation by Bacillus pumilus. Int J Biological Res 2: 44-47. |
| [58] | Gopi V, Upgade A, Soundararajan N (2012) Bioremediation potential of individual and consortium Non-adapted fungal strains on Azo dye containing textile effluent. Adv Appl Sci Res 3: 303-311. |
| [59] |
Ong ST, Keng PS, Lee WN, et al. (2011) Dye waste treatment. Water 3: 157-176. doi: 10.3390/w3010157
|
| [60] | Ratanapongleka K, Phetsom J (2014) Decolorization of synthetic dyes by crude laccase from Lentinus polychrous Lev. Int J Chem Eng Appli 5: 26-30. |
| [61] |
Bikovens O, Dizhbite T, Telysheva G (2012) Characterisation of humin substances formed during co-compositing of grass and wood wastes with animal grease. Environ Technol 33: 1427-1433. doi: 10.1080/09593330.2011.632652
|
| [62] |
Grinhut T, Hadar Y, Chen Y (2007) Degradation and transformation of humic substances by saprotrophic fungi: processes and mechanisms. Fungal Biol Rev 21: 179-189. doi: 10.1016/j.fbr.2007.09.003
|
| [63] | Mojsov K (2011) Application of enzymes in the textile industry: a review. II International Congress "engineering 230–239. |
| [64] | Gomathi VC, Ramanathan B, Sivaramaiah N, et al. (2012) Decolourization of paper mill effluent by immobilized cells of Phanerochaete chrysosporium. Int J Plant Anim Environ Sci 2: 141-146. |
| [65] |
Jadhav UP, Bholay AD, Shindikar M, et al. (2016) Bacterial lignin peroxidase mediated biobleaching and biodegradation of paper and pulp mill effluent. IOSR J Environ Sci Toxicol Food Technol 10: 31-36. doi: 10.9790/2402-1009023136
|
| [66] |
Andrady AL (2011) Microplastics in the marine environment. Mar Pollut Bull 62: 1596-1605. doi: 10.1016/j.marpolbul.2011.05.030
|
| [67] | Sowmya HV, Ramalingappa, Krishnappa M, et al. (2014) Biodegradation of polyethylene by Bacillus cereus. Adv Polymer Sci Technol Int J 4: 28-32. |
| [68] |
Wilkes RA, Aristilde L (2017) Degradation and metabolism of synthetic plastics and associated products by Pseudomonas sp.: capabilities and challenges. J Appl Microbiol 123: 582-593. doi: 10.1111/jam.13472
|
| [69] |
Roohi, Kulsoom B, Mohammed K, et al. (2017) Microbial enzymatic degradation of biodegradable plastics. Curr Pharm Biotechnol 18. doi: 10.2174/1389201018666170523165742
|
| [70] | Sharma M, Sharma P, Sharma A, et al. (2015) Microbial degradation of plastic-a brief review. CIBTech J Microbiol 4: 85-89. |
| [71] | Sowmya HV, Ramlingappa B, Krishnappa M, et al. (2014) Low density polyethylene degrading fungi isolated from local dumpsite of shivamogga district. Int J Biol Res 2: 39-43. |
| [72] |
Sowmya HV, Ramalingappa, Krishnappa M, et al. (2014) Degradation of polyethylene by Trichoderma harzianum--SEM, FTIR, and NMR analyses. Environ Monit Assess 186: 6577-6586. doi: 10.1007/s10661-014-3875-6
|
| [73] | Khatoon N, Jamal A, Ali MI (2018) Lignin peroxidase isoenzyme: a novel approach to biodegrade the toxic synthetic polymer waste. Environ Technol 40: 1-10. |
| [74] | Sowmya HV, Ramalingappa B, Nayanashree G, et al. (2015) Polyethylene degradation by fungal consortium. Int J Environ Sci Technol 9: 823-830. |
| [75] |
Krueger M, Harms H, Schlosser D (2015) Prospects for microbiological solution to environmental pollution with plastics. Appl Microbiol Biotechnol 99: 8857-8574. doi: 10.1007/s00253-015-6879-4
|
| [76] |
Dang T, Nguyen D, Thai H, et al. (2018) Plastic degradation by thermophilic Bacillus sp. BCBT21 isolated from composting agricultural residual in Vietnam. Adv Nat Sci Nanosci Nanotechnol 9: 015014. doi: 10.1088/2043-6254/aaabaf
|
| [77] | Gore S, Khotha A, Nevgi D, et al. (2017) The use of horse radish peroxidase, an eco-friendly method for removal of phenol from industrial effluent. J Environ Sci Toxicol Food Technol 11: 7-13. |
| [78] |
Shekoohiya S, Moussavi G, Naddafi K (2016) The peroxidase-mediated biodegradation of petroleum hydrocarbons in a H2O2-induced SBR using in-situ production of peroxidase: Biodegradation experiments and bacterial identification. J Hazard Mater 313: 170-178. doi: 10.1016/j.jhazmat.2016.03.081
|
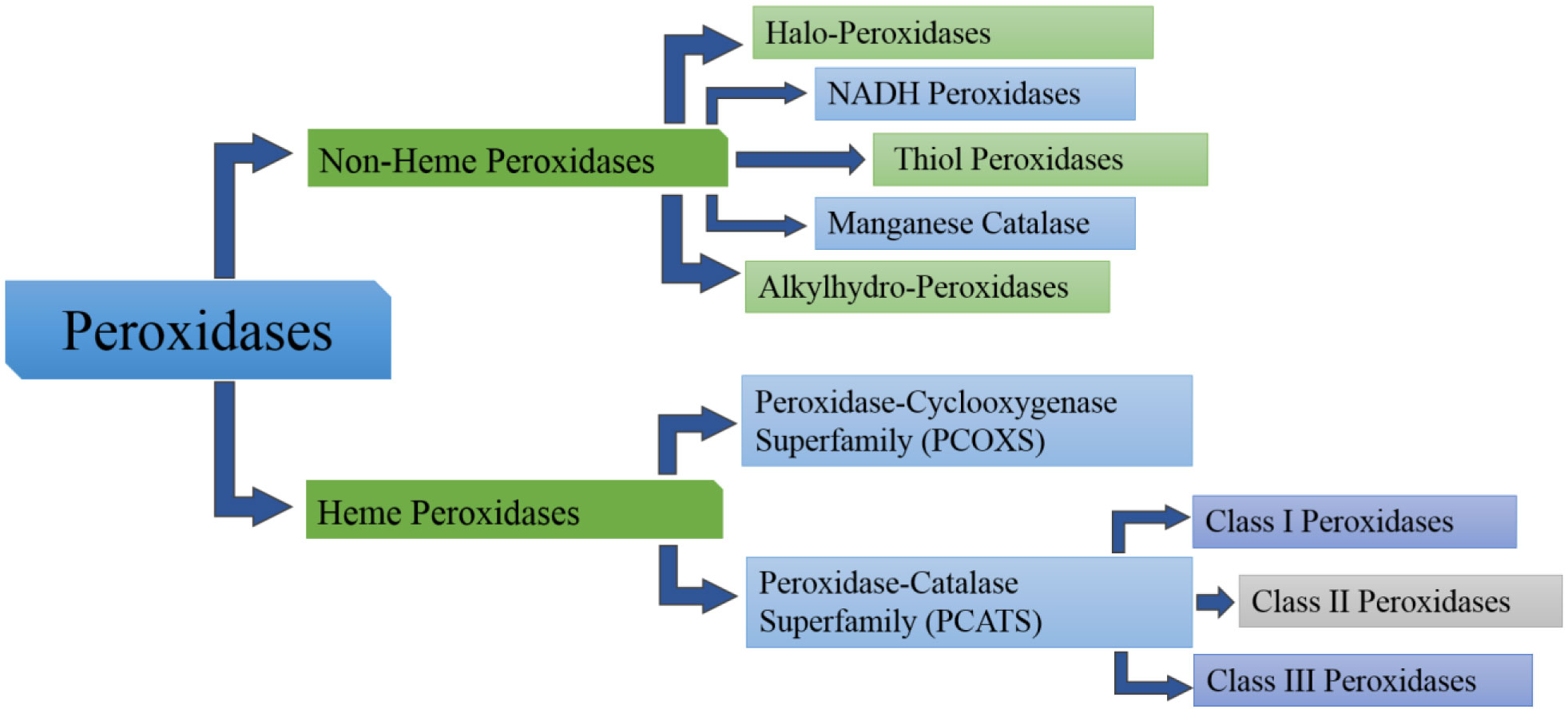
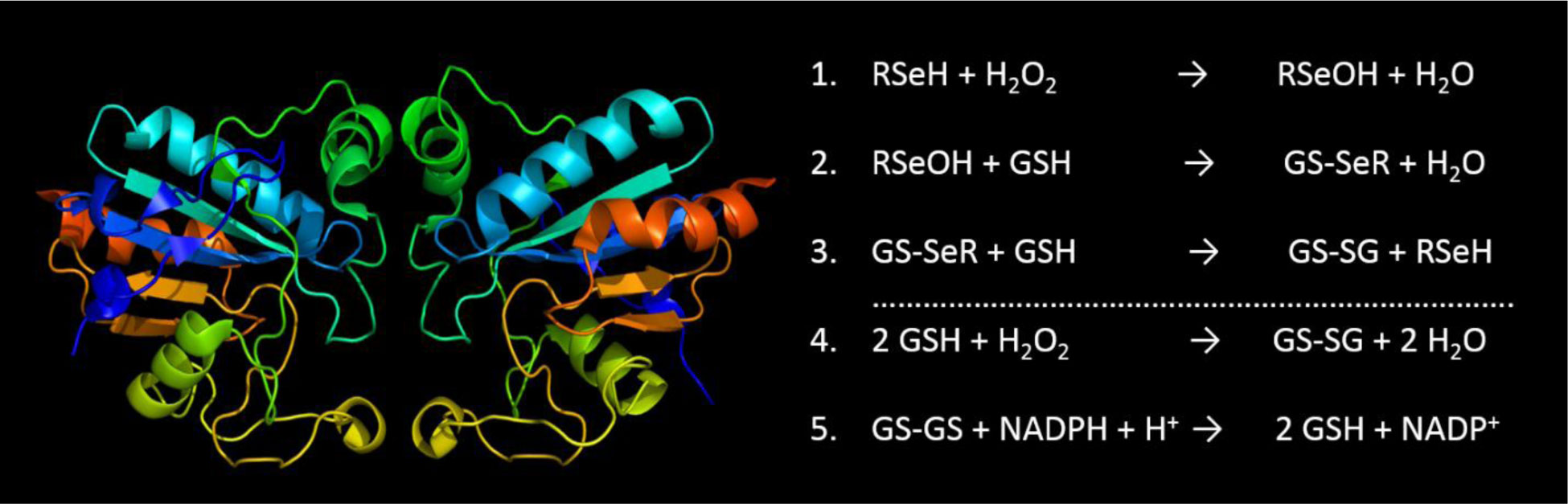
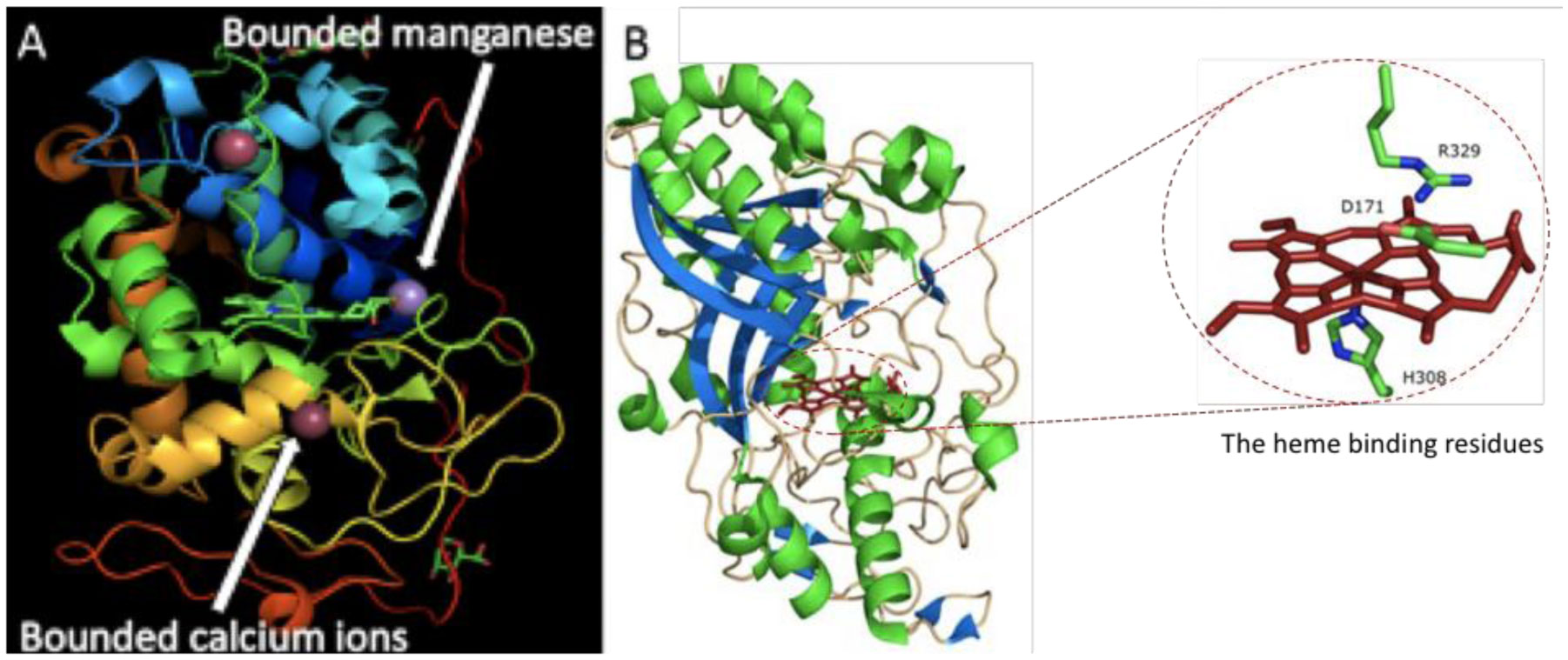
Figures(3) / Tables(1)

Pontsho Patricia Twala, Alfred Mitema, Cindy Baburam, Naser Aliye Feto. Breakthroughs in the discovery and use of different peroxidase isoforms of microbial origin[J]. AIMS Microbiology, 2020, 6(3): 330-349. doi: 10.3934/microbiol.2020020







 DownLoad:
DownLoad: