




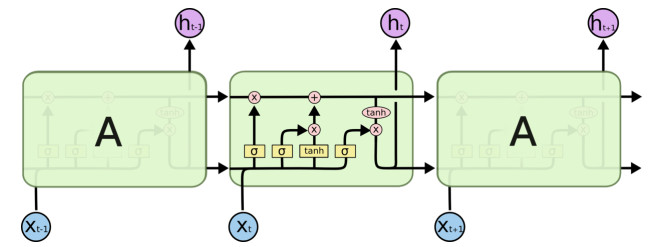
Figure 1.
The specific structure of a node of LSTM.
Citation: A. Burhan, G. Rasul, T. Qadir, S. Hussain, M. Saqib, S.A.A. Bukhari. Environmental policies to protect pollinators: attributes and actions needed to avert climate borne crisis of oil seed agriculture in Pakistan[J]. AIMS Agriculture and Food, 2017, 2(3): 233-250. doi: 10.3934/agrfood.2017.3.233
| [1] | Giuseppe Ciaburro . Machine fault detection methods based on machine learning algorithms: A review. Mathematical Biosciences and Engineering, 2022, 19(11): 11453-11490. doi: 10.3934/mbe.2022534 |
| [2] | Hanyu Zhao, Chao Che, Bo Jin, Xiaopeng Wei . A viral protein identifying framework based on temporal convolutional network. Mathematical Biosciences and Engineering, 2019, 16(3): 1709-1717. doi: 10.3934/mbe.2019081 |
| [3] | Chaofan Li, Kai Ma . Entity recognition of Chinese medical text based on multi-head self-attention combined with BILSTM-CRF. Mathematical Biosciences and Engineering, 2022, 19(3): 2206-2218. doi: 10.3934/mbe.2022103 |
| [4] | Darrak Moin Quddusi, Sandesh Athni Hiremath, Naim Bajcinca . Mutation prediction in the SARS-CoV-2 genome using attention-based neural machine translation. Mathematical Biosciences and Engineering, 2024, 21(5): 5996-6018. doi: 10.3934/mbe.2024264 |
| [5] | Chongyi Tian, Longlong Lin, Yi Yan, Ruiqi Wang, Fan Wang, Qingqing Chi . Photovoltaic power prediction based on dilated causal convolutional network and stacked LSTM. Mathematical Biosciences and Engineering, 2024, 21(1): 1167-1185. doi: 10.3934/mbe.2024049 |
| [6] | Lingyun Xiang, Guoqing Guo, Jingming Yu, Victor S. Sheng, Peng Yang . A convolutional neural network-based linguistic steganalysis for synonym substitution steganography. Mathematical Biosciences and Engineering, 2020, 17(2): 1041-1058. doi: 10.3934/mbe.2020055 |
| [7] | Chaofan Li, Qiong Liu, Kai Ma . DCCL: Dual-channel hybrid neural network combined with self-attention for text classification. Mathematical Biosciences and Engineering, 2023, 20(2): 1981-1992. doi: 10.3934/mbe.2023091 |
| [8] | Noor Wali Khan, Mohammed S. Alshehri, Muazzam A Khan, Sultan Almakdi, Naghmeh Moradpoor, Abdulwahab Alazeb, Safi Ullah, Naila Naz, Jawad Ahmad . A hybrid deep learning-based intrusion detection system for IoT networks. Mathematical Biosciences and Engineering, 2023, 20(8): 13491-13520. doi: 10.3934/mbe.2023602 |
| [9] | Jia-Gang Qiu, Yi Li, Hao-Qi Liu, Shuang Lin, Lei Pang, Gang Sun, Ying-Zhe Song . Research on motion recognition based on multi-dimensional sensing data and deep learning algorithms. Mathematical Biosciences and Engineering, 2023, 20(8): 14578-14595. doi: 10.3934/mbe.2023652 |
| [10] | Tao Yang, Qicheng Yang, Yibo Zhou, Chuanbiao Wen . Glucose trend prediction model based on improved wavelet transform and gated recurrent unit. Mathematical Biosciences and Engineering, 2023, 20(9): 17037-17056. doi: 10.3934/mbe.2023760 |
Information hiding, as an important research direction in the field of network security, has made great progress in recent years. Secret information is usually embedded into various multimedia carriers, such as text, images [1], audio [2] and videos [3]. Since text is the most widely used media, information hiding based on text has great value. Most traditional text steganography methods hide secret information in an existing document by exploiting its typesetting (format-based) and its lexical, syntactical, and semantical features (content-based) of the document. Format-based methods, for example, typically alter letter-spacing [4] and line-spacing [5], change the font attributes [6], and utilize invisible characters [7]. These methods, however, are sensitive to formats, and a slight change of format could cause new errors when extracting secret information. Common content-based methods, on the other hand, may change existing semantics using, e.g., synonym substitution [8,9], and adjust syntactic structures of certain sentences [10]. These traditional methods embed secret information by making some changes to the text, thus they are vulnerable to some steganalysis algorithms [11,12], which lead to poor security.
In recent years, researchers have started to investigate how to generate text, instead of modifying existing one, to hide information. For example, Yu et al. [13] proposed to use a special style of poetry called "Song Ci", originated in the Song Dynasty in ancient China, to form text steganography. This was the first text steganography algorithm to make use of a Chinese art form. However, their method only selects suitable words from an existing Song-Ci poem to recombines a new Ci-poem. It does not really generate new Ci-poems from scratch, and so may limit its usage. In addition, their method randomly chooses words during the generation process and ignores word collocations and the relationships between lines. Because of these, Ci-poems generated by their algorithms often lack a central theme, which might arouse suspicion and thus reduce security.
Recently, deep-learning technologies has achieved great success in the field of image generation [14]. Benefiting from this, more attempts of using such technologies have been made to generate a certain type of text form. For example, Zhang and Lapata [15] studied poem generations as a machine- translation problem using the standard encoder-decoder model. Their method compresses all previously generated lines for a poem to be generated into a vector to help generate the next line. Wang et al. [16,17] and Yi et al. [18] presented a method to generate a poem around a central theme using a bidirectional recurrent neural network (RNN) with attention mechanism, which allow them to focus on the most critical components and deep dependencies of lines in a poem.
Using CNN modeling, researchers have applied text generations to text steganography. For example, Luo et al. [19] presented a method based on Ci-poem generations using a Markov chain model. While their method does produce a better Ci-poem, it has a low embedding capacity for hiding information. They [20] also showed how to use a LSTM model to generate Chinese classic poems and embed secret information into a generated poem during the generation process.
We note that ancient styles of poetry, although once played an important role in the literature, often have a unique style of expressions that makes it difficult for general readers to comprehend. In particular, secret information embedded in a poem may look odd and thus arouse suspicion, leading to weaker security.
To overcome this obstacle, we devise a novel text steganography method using a RNN model to generate Chinese pop music lyrics. Compared to the poems written in a stringent style, lyrics are easier to understand and allowed to have a much longer length, yielding better security and a higher embedding capacity. However, if applying neural networks to lyrics generation directly, especially when combining text steganography, the resulting text may not look like lyrics at all. The reason is that lyrics must also follow a certain structure and properly rhyme. Moreover, lyrics should express emotions consistently. To tackle these issues, we analyze a large number of Chinese pop lyrics, summarize three common types, and use them as structure templates to guide lyric generation. We then present thirteen rhymes, similar to the oblique tones of poems, for generating lyrics with pleasant tones.
Our contributions are the following: First, we devise a RNN-based lyric generator with steganography to offer a much higher embedding capacity than previous steganography methods. Second, we present three structure templates for Chinese pop music and thirteen rhymes to mitigate the lyric-quality decrease incurred during the process of hiding information.
The rest of the paper is organized as follows: In Section 2 we describe preliminary results. In Section 3 we present a detailed description of our RNN-based lyric-generation model suitable for text steganography. In Section 4 we carry out the construction of dataset, show the generated results, evaluate the proposed method and make comparisons with previous methods. Finally, we conclude the paper in Section 5.
In this section, we describe our preliminary results on Char-RNN, lyric formats, and rhymes.
The sequence-to-sequence (S2S) model has been widely used in machine translation, automatic response, text generation, and other fields. S2S may be carried out under the framework of Encoder-Decoder and implemented as a RNN, CNN or GRU. Recurrent Neural Network (RNN) is the most widely used, which has been proved to be the most suitable network model for dealing with sequential tasks. However, a basic RNN model consists of only one hidden state, and the information at the beginning of the sequence is seldom retained in a later stage, there is a severe problem of gradient disappearance [17]. Therefore, basic RNNs are not suitable for generating the long lyrics. To solve these problems, researchers have proposed a number of variants of RNN, including LSTM and GRU.
As a special kind of RNN network, LSTM [21] adopts four processing methods that interact with each other in a special way to realize the processing of long-term dependence. The LSTM network model is shown in Figure 1. All recurrent neural networks have the form of a chain of repeating modules of neural network. In standard RNNs, this duplicate module has only one very simple structure, such as a single layer. However, LSTM avoids long-term dependency problems through special design. It usually uses a structure called a 'gate' to remove or increase the transmission of information.
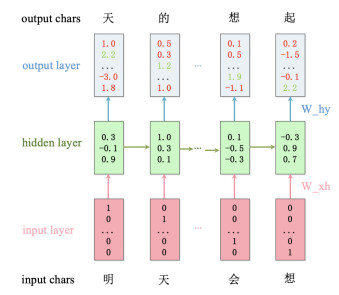
In 2014, Karpathy [22] proposed a Char-RNN model, allowing machines to generate text from the dimension of characters. In other words, the model computes the probability of the next character based on the observed ones. In this paper, we devise a Char-RNN model based on LSTM to deal with long-distance dependency problems, Figure 2 shows the execution process of the model.
The Chinese pop music lyrics follow certain formats. After analyzing a large number of them, we found that most of them follow the following three structures: (1) A1 + B, A2 + B, (2) A1 + A2 + A3, (3) A1 + A2 + B1 + B2, where each letter with or without a number represents a lyric component consisting of 3 to 5 lines. Figure 3 shows an example of a classic 4-segment lyric.

A good Chinese pop lyric expresses strong emotions that are properly rhymed. To make Chinese pop lyrics rhyme, lyric writers would need to make sure that the pronunciations of the end words of each line rhyme. From the perspective of vocal music, Chinese syllables are divided into thirteen categories, which are called Thirteen-Rhymes [23]. The thirteen rhymes are classified according to the basic principles of the same or similar vowels. Table 1 shows the thirteen rhymes and the common Chinese characters for each rhyme. Figure 4 shows the content of the entire lyric "Deskmate of You", which is popular in mainland China. We can see that the rhyme of this lyric is Yi-Qi (pronunciations). In addition, a lyric may have more than one rhyme, and it is common to have three to five rhymes in a lyric.
| Rhymes | Vowels | Characters | Music |
| Fa-Hua | a, ia, ua | 巴、麻、花、沙 | 《绒花》 |
| Suo-Bo | o, e, uo | 波、多、朵、烁 | 《我爱你中国》 |
| Ye-Xie | ie, ue, üe | 街、谐、月、夜 | 《娄山关》 |
| Gu-Su | u | 出、珠、福、图 | 《闪光的珍珠》 |
| Yi-Qi | i, ü, er | 溪、雨、曲、句 | 《同桌的你》 |
| Huai-Lai | ai, uai | 白、海、爱、来 | 《四季歌》 |
| Hui-Dui | ei, ui, uei | 飞、灰、梅、泪 | 《大森林的早晨》 |
| Yao-Tiao | ao, iao | 高、烧、苗、笑 | 《绣荷包》 |
| You-Qiu | ou, iu, iou | 沟、球、舟、柳 | 《蝶恋花》 |
| Yan-Qian | an, ian, uan, üan | 帆、天、旋、唤 | 《黄河颂》 |
| Ren-Chen | en, in, un, ün | 春、纷、新、云 | 《兰花花》 |
| Jiang-Yang | ang, iang, uang | 芳、霜、香、长 | 《松花江上》 |
| Zhong-Dong | eng, ing, ueng, ong | 风、星 | 《故乡是北京》 |
 DownLoad:
CSV
DownLoad:
CSV

Information hiding starts when the lyric generator produces a vector Y on the input vector X. We denote the output vector by Y=y1,y2,…,y3, where N is the length of input and output sequences. Moreover, yi(i=1,2,…,N−1) is equal to xi(i=2,3,…,N); in other words, YN is obtained from yi(i=1,2,...,N−1) with the highest probability and the model generates next y based on yi(i=2,3,...,N). Here YN could be a character or word. Thus, to hide secret information, we will filter out candidate characters or words on selected positions, code them, and select one of the most suitable candidates in accord with the secret information.
There are three steps: (1) Set parameters. (2) Generate candidates for the next character (or word). (3) Select the most suitable candidate. The whole process of information hiding is shown in Figure 5. In Step 1, we set the following four parameters:
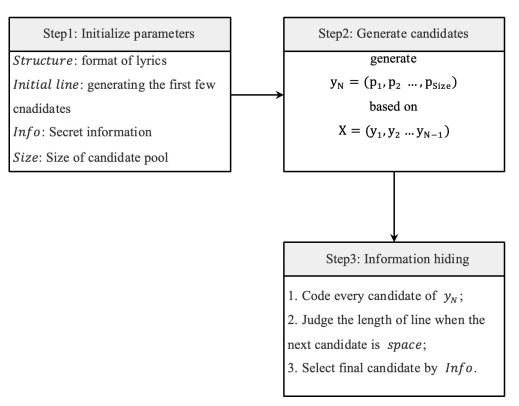
● Structure. This was the underlying structure for the lyric to be generated, which was described in Section 2.2.
● Initialline. It preheats the generator and helps to generate the first few characters, but it is not part of the final lyrics.
● Information. This is a binary stream converted from the secret information, which is hidden, transformed and extracted.
● Size. It is the size of a candidate pool, from which we choose the most probable candidates. It is obvious that is an important factor of the embedding capacity. The larger Size is, the more secret information can be embedded.
In Step 2, the lyric generator produces YN = (p1, p2, ..., pSize) based on (y1, y2, ..., yN−1), where pi=1,2,...,Size are the probability of Size candidates.
Step 3 is the most critical part of our lyric-steganography method. For each YN, we first predict all the possibilities of the next characters based on the previous information. We then select m most likely characters as candidates, where m = Size. We encode these m characters (such as Huffman code). Finally, we choose the most suitable character to hide the secret information. In addition, it should be pointed out that if YN is space, we need to judge whether the length of the sentence has reached the minimum requirement. If it is, then the sentence is finished. Otherwise, we need to change the order of the candidates so that lines do not end too early.
In order to improve the quality of generated lyrics, we applied lyrics structures and rhythm table to experiments, which mentioned above, and propose the Word-RNN model. Then, two results are showed, which are generated by two different models respectively. Finally, we illustrate the extraction method of secret information.
As described in Section 2.2, most Chinese pop lyrics follow three common structures. Thus, we introduce three structure templates to regulate the lyric generations. Taking Structure 3 as an example, we may generate four-verse lyrics with four sentences in each verse. It is common for a line in a verse to contain about 8 to 15 characters. To standardize the structure, it is crucial to control the number of characters per line. Therefore, in the process of creating lyrics, we will make a judgment on the number of characters in each line. If the number of characters has not reached the limit, no space will be generated. If the number of words exceeds the maximum, start a new line.
In addition, as mentioned in the Section 2.3, the ending character in each line should have proper rhyme. Therefore, during the generation process, we first determine whether the character to be generated is the ending character of the line. If it is, then we need to select the m characters with the highest probability from all the results, and these m characters rhyme properly. We then use the same method in Section 3.1 to choose the most suitable character for YN. If it were not the ending character, then we will choose m highest probable characters directly from all probability as the candidates.
Directly choosing the most suitable character may incur a severe problem, that is, the chosen character may not have a reasonable meaning with adjacent characters. Thus, the quality of lyric will decrease sharply. To overcome this problem, we improve the Char-RNN model to design a Word-RNN model, which have the same structure as Char-RNN. The only difference is that the smallest unit in the training and generation stage is a word instead of a character. Figure 6 shows the principle of Word-RNN.

We present an example to show how to hide the secret information in the process of lyric generations. First, we set the parameters as follows:
● Initial Line:我们手牵手(Your hands in my hands).
● Size: 4 (the size of candidate pool).
● Structure: Classical-16 (that is, four verses with four lines in each verse).
● Information: 100111111100001101.... (converted from '信息隐藏').
At the beginning, we choose a model from Char-RNN and Word-RNN (here we use Char-RNN as example). Then, the initial line will be fed to preheat the model and converted into xi.
For the first y, we select the most probable top-4 characters as candidates. We then encode the characters using Huffman code (shown in Table 2, Col. 1). When the beginning of the information is 10, then '你' is chosen as the first character.
| Bits | Char1 | Char2 | Char3 | Char4 | Char5 | Char6 | Char7 | Char8 | Char9 |
| 00 | 一 | 说 | 心 | space | 再 | 手 | 不 | 放 | space |
| 01 | 我 | 的 | 爱 | 心 | 会 | space | 我 | 再 | 留(歇) |
| 10 | 你 | 们 | 眼 | 我 | 在 | 开 | 的 | 要 | 地 |
| 11 | 这 | 要 | 手 | 不 | 放 | 不 | 让 | 停 | 的 |
DownLoad:
CSV
To make the ending characters of lines rhyme, we note that the bits of '01' represent character '歇' (shown in Table 2). However, this character does not sound well with the You-Qiu rhyme of the existing lines. Thus, we replace it with another character that rhyme better. Also, it should be noted that the space character in Table 2 is used as the separator between two lines, which can also be used to hide information.
After a lyric embedded with secret information is generated, the sender may send the lyric to the receiver. Meanwhile, the file about candidate pool should be sent together. When the receivers obtain the lyric, they can extract the binary stream converted from secret information according to the candidate pool. In other words, it is impossible to extract secret information without the file of candidate pool. Thus, the file of candidate pool plays an important role in the process of information extraction, because only the legitimate recipients can get the file and extract the secret information.
In this section, we first illustrate the construction of dataset and how to set the value for each initial parameter. Then, we present two results, which are generated by Char-RNN and Word-RNN respectively. Finally, the performance evaluations are given.
We need a large number of high-quality lyrics to train a satisfactory model. Unfortunately, no Chinese pop lyrics data has been published by any authoritative organization. To obtain training data, we use the Scrapy crawler framework to collect a total of 15,000 Chinese pop lyrics from music websites, including 100 Chinese male singers, 100 Chinese female singers, 100 Chinese bands, and 50 popular songs of each singer. Removing all-English songs and duplicates, we finally obtained a dataset of about 13,500 Chinese pop lyrics.
In the training process, we count all the Chinese characters in the lyric data, and construct a dictionary, to be shared by both inputs and outputs. At the same time, we count the number of Chinese characters as the length of One-Hot Encoding. Finally, we train our model using tensorflow.
For the Word-RNN model, we use the same methods, parameters, and optimization algorithm as the Char-RNN model. The only difference is that we first carry out word segmentation of the lyric data, expressing each song in terms of phrases. Namely, in the One-Hot Encoding, each position now represents a word, instead of a character.
Figure 7 depicts two Chinese pop lyrics generated by the proposed method with steganography, where (a) is a lyric generated by Char-RNN and the (b) is a lyric generated by Word-RNN.

According to their literal meanings, we can consider that Lyric (a) praises the beauty of love, and Lyric (b) describes sorrow of love. Through the analysis of lyrics, we can find that the lyrics generated by Char-RNN model contain repetitive characters (in red) and broken lines, while the lyrics generated by the Word-RNN model do not have these problems.
We evaluate the proposed steganography method from two important aspects: embedding capacity and security.
Embedding capacity is a significant measure to evaluate a steganogaphy methods, which represents how much information can hide in a stego-text. Previous steganography methods based on text generations either have low embedding capacities or do not look natural. For example, the poetry-based methods not only generate a poor naturalness genre, but also have lower embedding capacity than the proposed method. The high embedding capacity of the proposed method stems from longer length of lyrics, and we can hide information at each char-position or word-position. Similarly, we can conclude that char-based methods have higher embedding capacities than word-based methods when the generated lyrics length are the same. The embedding capacity calculation formula of two methods are as follows:
| EC1=(num∑i=1Ci+num−1)∗log2Size | (4.1) |
| EC2=(num∑i=1Wi+num−1)∗log2Size | (4.2) |
Where EC1 represents the embedding capacity of a lyric generated by Char-RNN, num is the number of lines, Ci is the number of characters in each line, and Size is the number of candidates. Likewise, EC2 represents the embedding capacity of a lyric generated by Word-RNN method, where Wi is the number of words in each line.
Figure 8 depicts the comparison results of embedding capacities among different methods, where the value of the embedding capacity is the number of characters embedded in the text carriers. It is easy to see that the proposed methods in this paper have higher embedding capacities than Poem-based steganography method and Ci-based steganography method, and with increase of Size, the capacity will be much higher than these two methods.
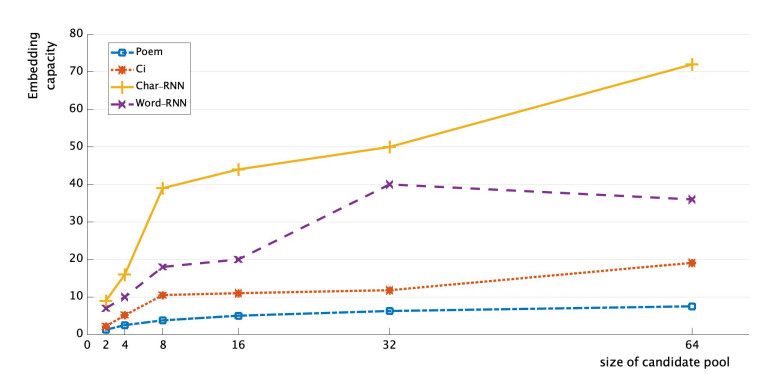
Security refers to the invisibility and imperceptibility of secret information, and it is one of the important measurements to evaluate a steganography method.
In modern society, people usually communicate with vernacular. Although poetry is an important achievement in the development of ancient Chinese literature, it is not practical in daily communication in modern society. Therefore, steganography based on poetry generation maybe easily arouses suspicion in the process of text carrier dissemination. Modern popular lyrics have higher naturalness, which means that the lyric text embedded with secret information will not arouses suspicion in the transmission process, thus the security is higher than steganography method based on poetry.
As shown in the Table 3, the results are generated by previous methods, which are embedded with secret information. Obviously, compared with the two results, the lyrics generated by the proposed method are easier to understand and has higher naturalness. Moreover, the proposed method is based on lyrics generation, which does not modify the text carrier like traditional text steganography methods, so it can effectively resist the detection of traditional steganalysis algorithms. Besides, our method is not based on text format, so it could resist the format-based attacks and steganalysis.
| Tang Poetry | Song Ci |
| 风窗烟树中, | 不知多少, 洞天谁道在, 一笑樽前. |
| 柔静雨光斑. | 底事今年春事早, 回首当日三贤. |
| 夜里吹生下, | 几度春风, 西风明月, 千里倍潸然. |
| 归人乱浪宽. | 断肠风月, 少年无限当年. |
DownLoad:
CSV
We presented a novel method that uses recurrent neural networks to generate the Chinese pop lyrics, and in the process of generation, we successfully embedded the secret information into the lyrics. We first used Char-RNN to predict the probabilities of the possible next characters based on the observed data. We then selected the most suitable character according to the secret information and added the character to the observed sequence, in order to continue to predict and generate. Directly generating lyrics based on secret information, it is likely that the generated characters can not be combined into words. We improved the Char-RNN model and developed the Word-RNN model, using words as units to train the model. Through experimental comparisons, we found that the proposed method offers higher hiding capacity and can generate more natural text, compared with the previous methods. In the future, we will improve our method to generate lyrics of better quality and clearer theme, with higher embedding capacity and better security and robustness.
This work was partially supported by National Natural Science Foundation of China (No. 61872134, 61502242), Natural Science Foundation of Hunan Province (No. 2018JJ2062, 2018JJ2301), and National Key Research and Development Program (2017YFC1703306), and Hunan Provincial 2011 Collaborative Innovation Center for Development and Utilization of Finance and Economics Big Data Property (2017TP1025).
The authors declare no conflict of interest.
| [1] |
Ollerton J, Rachael W, Sam T (2011) How many flowering plants are pollinated by animals? Oikos 120: 321-326. doi: 10.1111/j.1600-0706.2010.18644.x

|
| [2] |
Potts SG, Roberts SPM, Dean R, et al. (2010) Declines of managed honeybees and beekeepers in Europe? J Apic Res 49: 15-22. doi: 10.3896/IBRA.1.49.1.02
|
| [3] |
Gallai N, Jean MS, Josef S, et al. (2009) Economic valuation of the vulnerability of world agriculture confronted with pollinator decline. Ecol Econ 68: 810-821. doi: 10.1016/j.ecolecon.2008.06.014
|
| [4] |
Regan EC, Luca S, Lisa I, et al. (2015) Global trends in the status of bird and mammal pollinators. Conserv Lett 8: 397-403. doi: 10.1111/conl.12162
|
| [5] |
Eilers EJ, Kremen C, Smith GS, et al. (2011) Contribution of pollinator-mediated crops to nutrients in the human food supply. PLoS One 6: e21363. doi: 10.1371/journal.pone.0021363
|
| [6] | IPCC (2007) Climate change 2007: synthesis report - contribution of Working Groups 1, 2 and 3 to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. In: Change IPoC, Geneva. |
| [7] |
Memmott J, Craze PG, Waser NM, et al. (2007) Global warming and the disruption of plant-pollinator interactions. Ecol Lett 10: 710-717. doi: 10.1111/j.1461-0248.2007.01061.x
|
| [8] | Karim K. Climate Change Cost Pakistan 3.5 Billion Dollars in 18 Years. 2009. Available from: http://freebird.instablogs.com/entry/climate-change-cost-pakistan-35-billion-in-18years/#ixzz0qnHzBhRM |
| [9] | Schweiger O, Biesmeijer JC, Bommarco R, et al. (2010) Multiple stressors on biotic interactions: how climate change and alien species interact to affect pollination. Biol Rev 85: 777-795. |
| [10] |
Van Der Putten WH, De Ruiter PC, Bezemer TM, et al. (2004) Trophic interactions in a changing world. Basic Appl Ecol 5: 487-494. doi: 10.1016/j.baae.2004.09.003
|
| [11] |
Sutherst RW, Maywald GF, Bourne AS (2007) Including species interactions in risk assessments for global change. Glob Change Biol 13: 1843-1859. doi: 10.1111/j.1365-2486.2007.01396.x
|
| [12] | Oliver TH, Isaac NJB, August TA, et al. (2015) Declining resilience of ecosystem functions under biodiversity loss. Nat Commun 6: 2041-1723. |
| [13] | UN-Habitat Cities and Climate Change Initiative (2014) e-Newsletter. |
| [14] | LWEC. Improving urban grassland for people and wildlife. Living With Environmental Change Policy and Practice Notes Note. 2016, Available from: http://www.nerc.ac.uk/research/partnerships/ride/lwec/ppn/ppn32/ |
| [15] | Dicks LV, Mathilde B, Stuart PMR, et al. (2015) How much flower-rich habitat is enough for wild pollinators? Answering a key policy question with incomplete knowledge. Ecol Entomol 40: 22-35. |
| [16] |
Santangeli A, Lynn VD, Irina H, et al. (2016) Voluntary non-monetary approaches for implementing conservation. Biol Conserv 197: 209-214. doi: 10.1016/j.biocon.2016.03.013
|
| [17] |
Parmesan C (2006) Ecological and evolutionary responses to recent climate change. Annu Rev Ecol Evol Syst 37: 637-669. doi: 10.1146/annurev.ecolsys.37.091305.110100
|
| [18] | Suttle KB, Thomsen MA, Power ME (2007) Species interactions reverse grassland responses to changing climate. Science 80: 640-642. |
| [19] |
Walther GR (2010) Community and ecosystem responses to recent climate change. Philos Trans R Soc B Biol Sci 365: 2019-2024. doi: 10.1098/rstb.2010.0021
|
| [20] | IPCC Climate Change (2014) Impacts, Adaptation, and Vulnerability Working Group II Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Camb Univ Press. |
| [21] | CBD COP6. Retired sections: paragraphs 4-6, 14-15, 17 and 21 Agricultural biological diversity Element 3. Capacity-building. 2002. Available from: www.cbd.int/decision/cop/?id=7179 |
| [22] | Rao GM, Suryanarayana MC, Thakar CV (1980) Bees can boost oilseed production. Indian Farming 29: 25-26. |
| [23] | Free JB (1993) Insect pollination of crops, 2nd edn. Academic, Harcourt Brace Jovanovich Publishing London. 684. |
| [24] | Abrol DP (2007) Foraging behaviour of Apis mellifera and Apis cerana as determined by the energetics of nectar production in different cultivars of Brassica campestris var. toria. J Apic Sci 51: 5-10. |
| [25] | Abrol DP (2008) Plant insect interaction in crucifers. In: Gupta SK (ed) Biology and breeding of crucifers. CRC Press/Taylor and Francis USA. 129-149. |
| [26] | Abrol DP (2009) Bees and beekeeping in India, 2nd edn. Kalyani Publishers, New Delhi, 720. |
| [27] | Mishra RC, Kumar J, Gupta JK (1988) The effect of mode of pollination on yield and oil potential of Brassica campestris L. var. sarson with observation on insect pollinators. J Apic Res 27: 186-189. |
| [28] | Kluser S, Peduzzi P (2007) Global pollinator decline: A literature review. UNEP/GRID, Geneva. |
| [29] |
Potts SG, Biesmeijer JC, Kremen C, et al. (2010) Global pollinator declines: Trends, impacts and drivers. Trends Ecol Evol 25: 345-353. doi: 10.1016/j.tree.2010.01.007
|
| [30] |
Cleland EE, Chuine I, Menzel A, et al. (2007) Shifting plant phenology in response to global change. Trends Ecol Evol 22: 357-365. doi: 10.1016/j.tree.2007.04.003
|
| [31] |
Kudo G, Nishikawa Y, Kasagi T, et al. (2004) Does seed production of spring ephemerals decrease when spring comes early? Ecol Res 19: 255-259. doi: 10.1111/j.1440-1703.2003.00630.x
|
| [32] | Deutsch CA, Tewksbury JJ, Huey RB, et al. (2008) Impacts of climate warming on terrestrial ectotherms across latitude. Proc Nat Acad Sci U S A 105: 6668-6672. |
| [33] | Zahedi H, Shirani RAH, Noor MG, et al. (2009) The effects of zeolite soil application and selenium foliar applications on growth yield and yield components of three canola cultivars under drought stress. World Appl Sci J 7: 255-262. |
| [34] |
Kimball BA (1983) Carbon dioxide and agricultural yield: An assemblage and analysis of 430 prior observations. Agron J 75: 779-788. doi: 10.2134/agronj1983.00021962007500050014x
|
| [35] |
Griffin KL, Seemann JR (1996) Plants, CO2 and photosynthesis in the 21st century. Chem Biol 3: 245-254. doi: 10.1016/S1074-5521(96)90104-0
|
| [36] |
Taub DR, Miller B, Allen H (2008) Effects of elevated CO2 on the protein concentration of food crops: A meta-analysis. Glob Change Biol 14: 565-575. doi: 10.1111/j.1365-2486.2007.01511.x
|
| [37] |
Harris HC, McWilliams JR, Mason WK (1978) Influence of temperature on oil content and composition of sunflower. Aust J Agric Res 29: 1203-1212. doi: 10.1071/AR9781203
|
| [38] | Berthier E, Schiefer E, Clarke GKC, et al. (2010) Contribution of Alaskan glaciers to sea-level rise derived from satellite imagery, Nat Geosci 3: 92-95. |
| [39] |
Meier MF, Dyurgerov MB, Rick UK, et al. (2007) Glaciers dominate eustatic sea-level rise in the 21st century. Science 317: 1064. doi: 10.1126/science.1143906
|
| [40] |
Hegland SJ, Nielsen A, Lázaro A, et al. (2009) How does climate warming affect plant-pollinator interactions? Ecol Lett 12: 184-195. doi: 10.1111/j.1461-0248.2008.01269.x
|
| [41] |
Cane JH (2002) Pollinating bees (Hymenoptera: Apiformes) of US alfalfa compared for rates of pod and seed set. J Econ Entomol 95: 22-27. doi: 10.1603/0022-0493-95.1.22
|
| [42] |
Inouye DW (2008) Effects of climate change on phenology, frost damage, and floral abundance of montane wildflowers. Ecology 89: 353-362. doi: 10.1890/06-2128.1
|
| [43] |
Ravi S, James DC, Alex F, et al. (2013) Rolling with the flow: Bumblebees flying in unsteady wakes. J Exp Biol 216: 4299-4309. doi: 10.1242/jeb.090845
|
| [44] | Graham JM (1992) The Hive and the Honey Bee. Dadant and Sons, Hamilton, IL. |
| [45] | Polito VS, Warren CM, Dale EK (1996) Bud Development, Pollination, and Fertilization. Almond Prod Man Uni CA Agric Nat Res 1996: 98-102. |
| [46] | Thorp RW (1996) Bee Management for Pollination. Almond Prod Man Uni CA Agri Nat Res 1996: 132-138. |
| [47] |
Timmons AM, O'Brien ET, Charters YM, et al. (1995) Assessing the risks of wind pollination from fields of brassica napus ssp. oleifera. Euphytica 85: 417-423. doi: 10.1007/BF00023975
|
| [48] | Thompson CE, Squire G, Mackay GR, et al. (1999) Regional patterns of gene flow and its consequences for GM oilseed rape. Monogr-Br Crop Prot Counc 1999: 95-100. |
| [49] | Puškadija Z, Štefanić E, Mijić A, et al. (2007) Influence of weather conditions on honey bee visits (apis mellifera carnica) during sunflower (helianthus annuus l.) blooming period. Poljoprivreda 13: 230-233. |
| [50] | Gliddon C, Boudry P, Walker S (1999) Gene flow - a review of experimental evidence. Environ Impact Gene Modif Crops 1999: 65-79. |
| [51] | Feehan J, Harley M, Minnen J (2009) Climate change in Europe. Impact on terrestrial ecosystems and biodiversity. A review. Agron Sustain Dev 29: 409-421. |
| [52] | Wheeler TR, Craufurd PQ, Ellis RH, et al. (2000) Temperature variability and the yield of annual crops. Agric Ecosyst Environ 82: 159-167. |
| [53] |
Galloway LF, Cirigliano T, Gremski K (2002) The contribution of display size and dichogamy to potential geitonogamy in Campanula americana. Int J Plant Sci 163: 133-139. doi: 10.1086/324556
|
| [54] |
Pacini E, Nepi M, Vesprini JL (2003) Nectar biodiversity: A short review. Plant Syst Evol 238: 7-21. doi: 10.1007/s00606-002-0277-y
|
| [55] |
Mitchell RJ, Karron JD, Holmquist KG, et al. (2004) The influence of Mimulus ringens floral display size on pollinator visitation patterns. Funct Ecol 18: 116-124. doi: 10.1111/j.1365-2435.2004.00812.x
|
| [56] |
Hegland SJ, Totland O (2005) Relationships between species' floral traits and pollinator visitation in a temperate grassland. Oecologia 145: 586-594. doi: 10.1007/s00442-005-0165-6
|
| [57] |
Philipp M, Hansen T (2000) The influence of plant and corolla size on pollen deposition and seed set in Geranium sanguineum (Geraniaceae). Nord J Bot 20: 129-140. doi: 10.1111/j.1756-1051.2000.tb01556.x
|
| [58] |
Kudo G, Harder LD (2005) Floral and inflorescence effects on variation in pollen removal and seed production among six legume species. Funct Ecol 19: 245-254. doi: 10.1111/j.1365-2435.2005.00961.x
|
| [59] |
Akhalkatsi M, Lösch R (2005). Water limitation effect on seed development and germination in Trigonella coerulea (Fabaceae). Flora 200: 493-501. doi: 10.1016/j.flora.2005.06.001
|
| [60] |
Melser C, Klinkhamer PGL (2001) Selective seed abortion increases offspring survival in Cynoglossum officinale (Boraginaceae). Am J Bot 88: 1033-1040. doi: 10.2307/2657085
|
| [61] | Boyer JS, Westgate ME (2003) Grain yields with limited water. In: International conference on water-saving agriculture and sustainable use of water and land resources in arid and semi-arid areas: 2385-2394. Oxford, UK, Oxford Univ. Press. |
| [62] | Pakistan Bureau of Statistics, Pakistan Oilseed Development Board (PODB). |
| [63] | IPCC Climate Change (2014) Mitigation of Climate Change Working Group III Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Camb Univ Press. |
| 1. | Stephen Bondy, Arezoo Campbell, Water Quality and Brain Function, 2017, 15, 1660-4601, 2, 10.3390/ijerph15010002 | |
| 2. | Simeon Kayowa Olatunde, Abiodun Ayanfemi Ayandele, Microbiological and physico-chemical analyses of hand dug well-water near pit latrine in a rural Area of Western Nigeria, 2018, 12, 1996-0786, 132, 10.5897/AJEST2017.2463 | |
| 3. | Jin-Jing Xiao, Yang Li, Qing-Kui Fang, Yan-Hong Shi, Min Liao, Xiang-Wei Wu, Ri-Mao Hua, Hai-Qun Cao, Factors Affecting Transfer of Pyrethroid Residues from Herbal Teas to Infusion and Influence of Physicochemical Properties of Pesticides, 2017, 14, 1660-4601, 1157, 10.3390/ijerph14101157 | |
| 4. | Margaret Eggers, John Doyle, Myra Lefthand, Sara Young, Anita Moore-Nall, Larry Kindness, Roberta Other Medicine, Timothy Ford, Eric Dietrich, Albert Parker, Joseph Hoover, Anne Camper, Community Engaged Cumulative Risk Assessment of Exposure to Inorganic Well Water Contaminants, Crow Reservation, Montana, 2018, 15, 1660-4601, 76, 10.3390/ijerph15010076 | |
| 5. | Zijian Li, Health and safety assessment and regulatory management of Aldicarb, Atrazine, Diuron, Glyphosate, and MCPA by theoretical maximum daily intake estimation, 2018, 25, 18715532, 3, 10.1016/j.jchas.2017.09.003 | |
| 6. | Eid Brima, Toxic Elements in Different Medicinal Plants and the Impact on Human Health, 2017, 14, 1660-4601, 1209, 10.3390/ijerph14101209 | |
| 7. | Motoyuki Kamata, Yoshihiko Matsui, Mari Asami, National trends in pesticides in drinking water and water sources in Japan, 2020, 744, 00489697, 140930, 10.1016/j.scitotenv.2020.140930 | |
| 8. | Ibrahim Shabbaj, Mansour Alghamdi, Magdy Shamy, Salwa Hassan, Musaab Alsharif, Mamdouh Khoder, Risk Assessment and Implication of Human Exposure to Road Dust Heavy Metals in Jeddah, Saudi Arabia, 2017, 15, 1660-4601, 36, 10.3390/ijerph15010036 | |
| 9. | Young-seoub Hong, Byeong-jin Ye, Yu-mi Kim, Byoung-gwon Kim, Gyeong-hui Kang, Jeong-jin Kim, Ki-hoon Song, Young-hun Kim, Jeong-wook Seo, Investigation of Health Effects According to the Exposure of Low Concentration Arsenic Contaminated Ground Water, 2017, 14, 1660-4601, 1461, 10.3390/ijerph14121461 | |
| 10. | Zijian Li, Evaluation of regulatory variation and theoretical health risk for pesticide maximum residue limits in food, 2018, 219, 03014797, 153, 10.1016/j.jenvman.2018.04.067 | |
| 11. | Zijian Li, Aaron Jennings, Global variations in pesticide regulations and health risk assessment of maximum concentration levels in drinking water, 2018, 212, 03014797, 384, 10.1016/j.jenvman.2017.12.083 | |
| 12. | Shalini Sanyal, Sujata Law, Chronic pesticide exposure induced aberrant Notch signalling along the visual pathway in a murine model, 2021, 282, 02697491, 117077, 10.1016/j.envpol.2021.117077 | |
| 13. | Allan Pretti Ogura, Jacqueline Zanin Lima, Jéssica Pelinsom Marques, Lucas Massaro Sousa, Valéria Guimarães Silvestre Rodrigues, Evaldo Luiz Gaeta Espíndola, A review of pesticides sorption in biochar from maize, rice, and wheat residues: Current status and challenges for soil application, 2021, 300, 03014797, 113753, 10.1016/j.jenvman.2021.113753 | |
| 14. | Karina Bano, Sandeep Kaushal, Prit Pal Singh, A review on photocatalytic degradation of hazardous pesticides using heterojunctions, 2021, 209, 02775387, 115465, 10.1016/j.poly.2021.115465 | |
| 15. | Ke Fang, Yu‐Xin Xiang, Han Wang, Ming‐Kun Li, Si‐Yuan Jiang, Chen‐Jun Liu, Xin Yang, Su‐Wan Wei, Jin‐Jing Xiao, Yan‐Hong Shi, Hai‐Qun Cao, In vitro inhalation bioaccessibility and health risk assessment of difenoconazole in the atmosphere, 2023, 1526-498X, 10.1002/ps.7811 | |
| 16. | Anjana Bhardwaj, Preeti Yadav, Rajendra Vishwakarma, 2024, 9780443156601, 453, 10.1016/B978-0-443-15660-1.00018-6 |
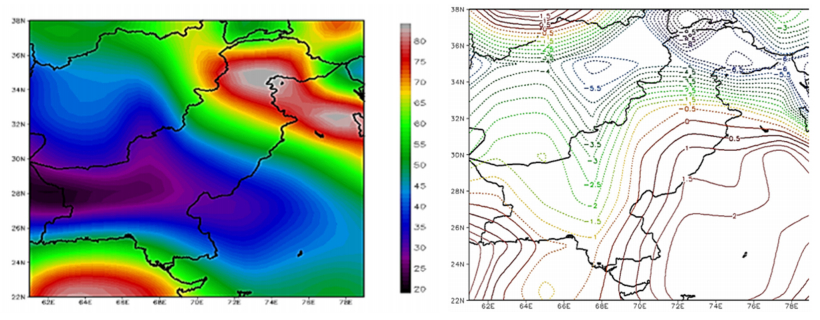
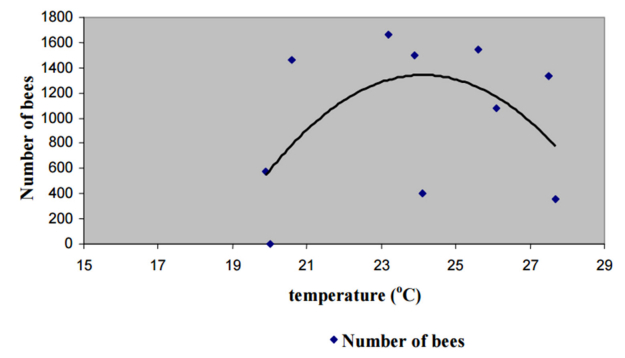
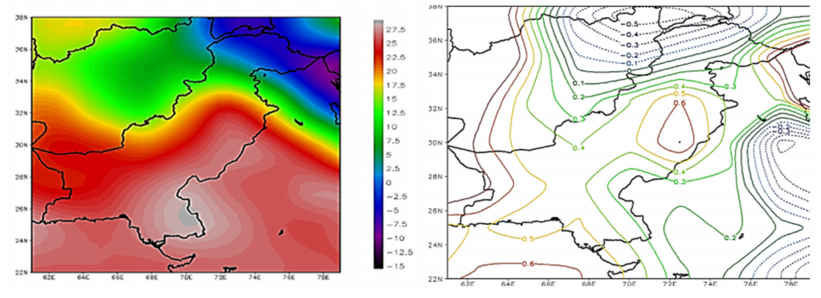
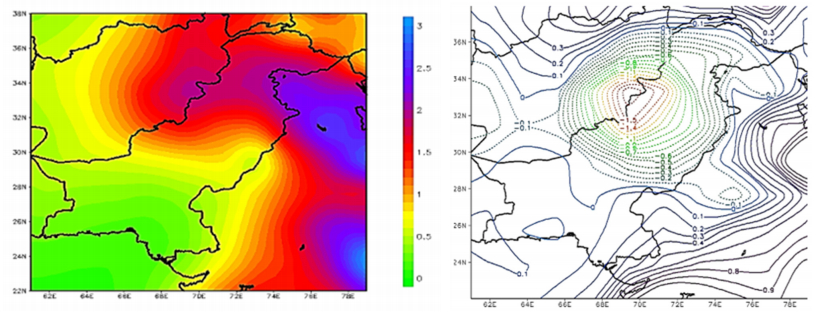
Figures(9) / Tables(1)

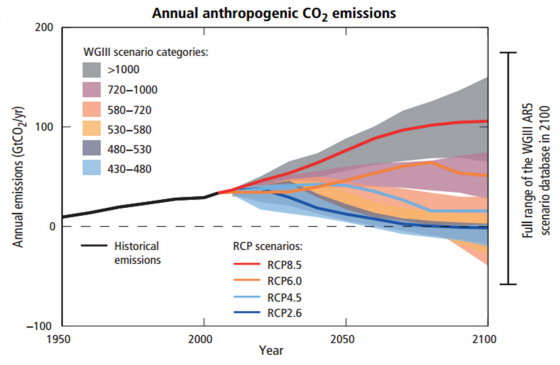
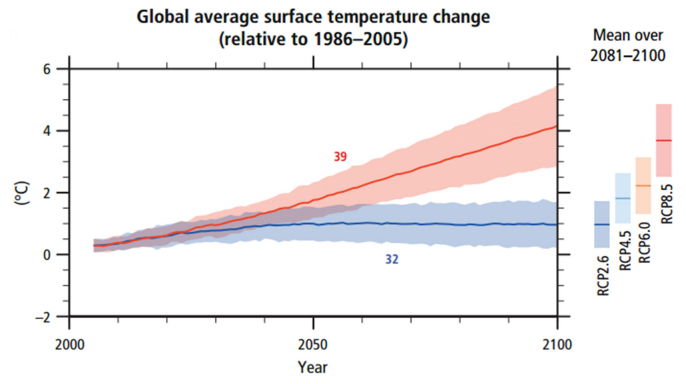
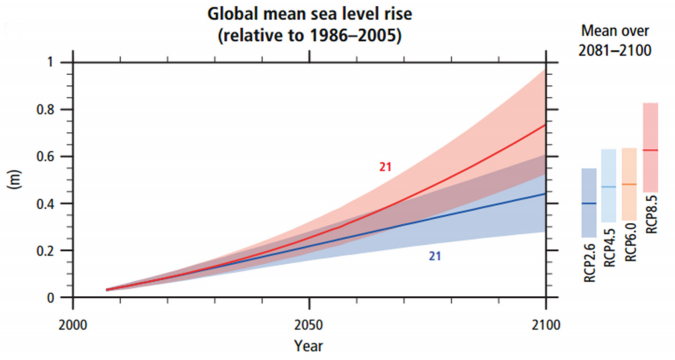
A. Burhan, G. Rasul, T. Qadir, S. Hussain, M. Saqib, S.A.A. Bukhari. Environmental policies to protect pollinators: attributes and actions needed to avert climate borne crisis of oil seed agriculture in Pakistan[J]. AIMS Agriculture and Food, 2017, 2(3): 233-250. doi: 10.3934/agrfood.2017.3.233

| Rhymes | Vowels | Characters | Music |
| Fa-Hua | a, ia, ua | 巴、麻、花、沙 | 《绒花》 |
| Suo-Bo | o, e, uo | 波、多、朵、烁 | 《我爱你中国》 |
| Ye-Xie | ie, ue, üe | 街、谐、月、夜 | 《娄山关》 |
| Gu-Su | u | 出、珠、福、图 | 《闪光的珍珠》 |
| Yi-Qi | i, ü, er | 溪、雨、曲、句 | 《同桌的你》 |
| Huai-Lai | ai, uai | 白、海、爱、来 | 《四季歌》 |
| Hui-Dui | ei, ui, uei | 飞、灰、梅、泪 | 《大森林的早晨》 |
| Yao-Tiao | ao, iao | 高、烧、苗、笑 | 《绣荷包》 |
| You-Qiu | ou, iu, iou | 沟、球、舟、柳 | 《蝶恋花》 |
| Yan-Qian | an, ian, uan, üan | 帆、天、旋、唤 | 《黄河颂》 |
| Ren-Chen | en, in, un, ün | 春、纷、新、云 | 《兰花花》 |
| Jiang-Yang | ang, iang, uang | 芳、霜、香、长 | 《松花江上》 |
| Zhong-Dong | eng, ing, ueng, ong | 风、星 | 《故乡是北京》 |
DownLoad:
CSV
| Bits | Char1 | Char2 | Char3 | Char4 | Char5 | Char6 | Char7 | Char8 | Char9 |
| 00 | 一 | 说 | 心 | space | 再 | 手 | 不 | 放 | space |
| 01 | 我 | 的 | 爱 | 心 | 会 | space | 我 | 再 | 留(歇) |
| 10 | 你 | 们 | 眼 | 我 | 在 | 开 | 的 | 要 | 地 |
| 11 | 这 | 要 | 手 | 不 | 放 | 不 | 让 | 停 | 的 |
DownLoad:
CSV
| Tang Poetry | Song Ci |
| 风窗烟树中, | 不知多少, 洞天谁道在, 一笑樽前. |
| 柔静雨光斑. | 底事今年春事早, 回首当日三贤. |
| 夜里吹生下, | 几度春风, 西风明月, 千里倍潸然. |
| 归人乱浪宽. | 断肠风月, 少年无限当年. |
DownLoad:
CSV
| Rhymes | Vowels | Characters | Music |
| Fa-Hua | a, ia, ua | 巴、麻、花、沙 | 《绒花》 |
| Suo-Bo | o, e, uo | 波、多、朵、烁 | 《我爱你中国》 |
| Ye-Xie | ie, ue, üe | 街、谐、月、夜 | 《娄山关》 |
| Gu-Su | u | 出、珠、福、图 | 《闪光的珍珠》 |
| Yi-Qi | i, ü, er | 溪、雨、曲、句 | 《同桌的你》 |
| Huai-Lai | ai, uai | 白、海、爱、来 | 《四季歌》 |
| Hui-Dui | ei, ui, uei | 飞、灰、梅、泪 | 《大森林的早晨》 |
| Yao-Tiao | ao, iao | 高、烧、苗、笑 | 《绣荷包》 |
| You-Qiu | ou, iu, iou | 沟、球、舟、柳 | 《蝶恋花》 |
| Yan-Qian | an, ian, uan, üan | 帆、天、旋、唤 | 《黄河颂》 |
| Ren-Chen | en, in, un, ün | 春、纷、新、云 | 《兰花花》 |
| Jiang-Yang | ang, iang, uang | 芳、霜、香、长 | 《松花江上》 |
| Zhong-Dong | eng, ing, ueng, ong | 风、星 | 《故乡是北京》 |
| Bits | Char1 | Char2 | Char3 | Char4 | Char5 | Char6 | Char7 | Char8 | Char9 |
| 00 | 一 | 说 | 心 | space | 再 | 手 | 不 | 放 | space |
| 01 | 我 | 的 | 爱 | 心 | 会 | space | 我 | 再 | 留(歇) |
| 10 | 你 | 们 | 眼 | 我 | 在 | 开 | 的 | 要 | 地 |
| 11 | 这 | 要 | 手 | 不 | 放 | 不 | 让 | 停 | 的 |
| Tang Poetry | Song Ci |
| 风窗烟树中, | 不知多少, 洞天谁道在, 一笑樽前. |
| 柔静雨光斑. | 底事今年春事早, 回首当日三贤. |
| 夜里吹生下, | 几度春风, 西风明月, 千里倍潸然. |
| 归人乱浪宽. | 断肠风月, 少年无限当年. |
