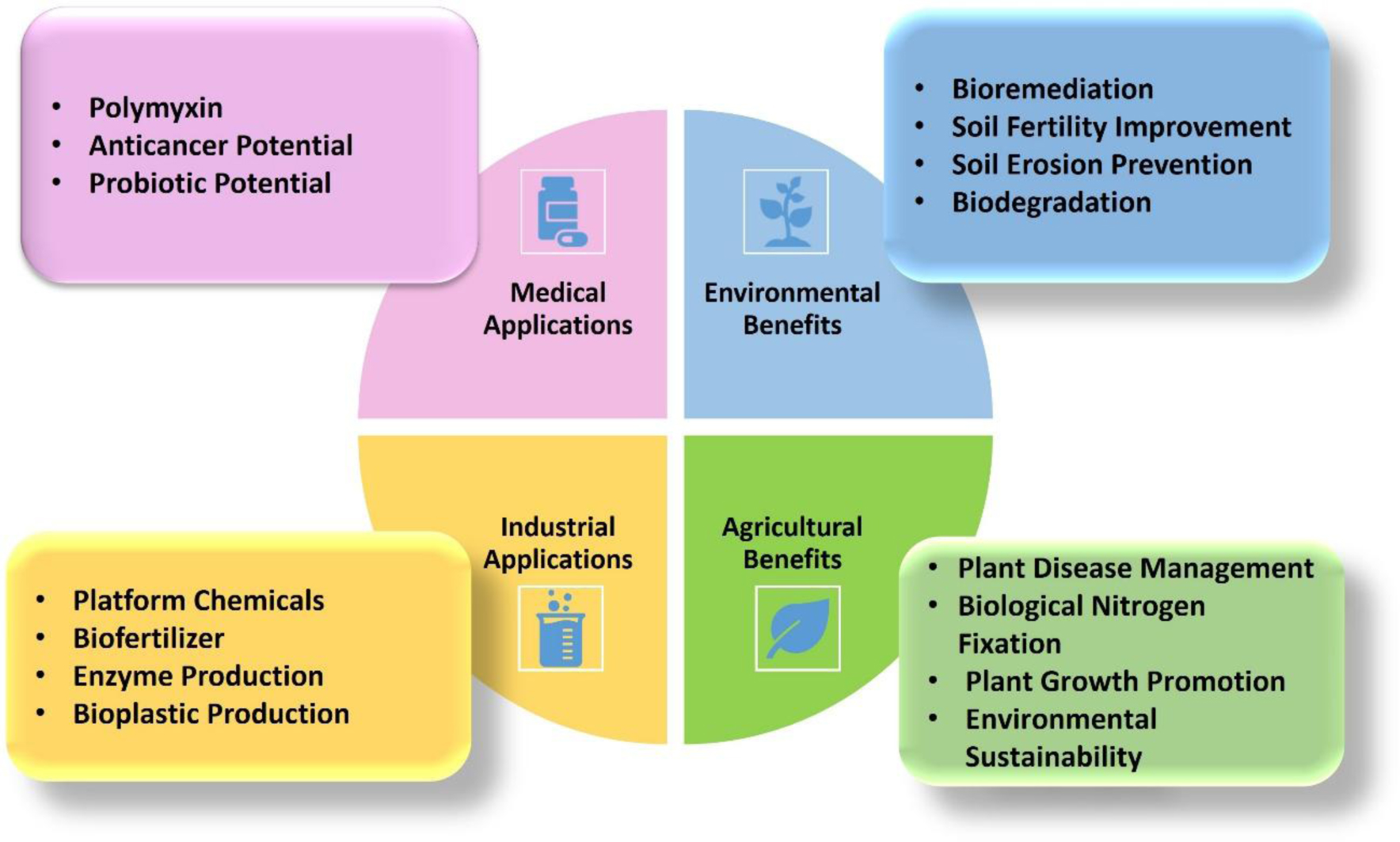
Paenibacillus polymyxa is a multifaceted bacterium with widespread applications in agriculture, environmental management, medicine, and industry. In agricultural settings, it plays a crucial role in soil enhancement, plant growth promotion, and natural pathogen control, reducing the need for chemical interventions. Additionally, P. polymyxa exhibits promising potential in medical applications by aiding in infection prevention and supporting gastrointestinal health. In the realm of environmental management, this bacterium contributes to pollution remediation through biodegradation processes. Industrially, P. polymyxa is involved in producing enzymes, biofertilizers, bioplastics, and platform chemicals, offering sustainable alternatives that underscore its importance in driving sustainability initiatives. Despite these valuable attributes, widespread utilization of bioresources derived from naturally occurring P. polymyxa has been hampered by limited genetic manipulation capabilities and tools. In this comprehensive analysis, we aimed to provide a thorough understanding of P. polymyxa's characteristics, genetic resources, and metabolic capabilities, while highlighting its potential as a versatile platform for protein expression, metabolic engineering, and synthetic biology. We delved into the diverse sustainable applications of P. polymyxa in these domains, emphasizing its benefits, challenges, and future outlook in advancing sustainable practices. Furthermore, we underscore the critical need for continued research and development of advanced engineering techniques and genetic editing technologies tailored specifically for this bacterium.
Citation: Imen Zalila-Kolsi, Ray Al-Barazie. Advancing sustainable practices with Paenibacillus polymyxa: From soil health to medical applications and molecular engineering[J]. AIMS Microbiology, 2025, 11(2): 338-368. doi: 10.3934/microbiol.2025016
Paenibacillus polymyxa is a multifaceted bacterium with widespread applications in agriculture, environmental management, medicine, and industry. In agricultural settings, it plays a crucial role in soil enhancement, plant growth promotion, and natural pathogen control, reducing the need for chemical interventions. Additionally, P. polymyxa exhibits promising potential in medical applications by aiding in infection prevention and supporting gastrointestinal health. In the realm of environmental management, this bacterium contributes to pollution remediation through biodegradation processes. Industrially, P. polymyxa is involved in producing enzymes, biofertilizers, bioplastics, and platform chemicals, offering sustainable alternatives that underscore its importance in driving sustainability initiatives. Despite these valuable attributes, widespread utilization of bioresources derived from naturally occurring P. polymyxa has been hampered by limited genetic manipulation capabilities and tools. In this comprehensive analysis, we aimed to provide a thorough understanding of P. polymyxa's characteristics, genetic resources, and metabolic capabilities, while highlighting its potential as a versatile platform for protein expression, metabolic engineering, and synthetic biology. We delved into the diverse sustainable applications of P. polymyxa in these domains, emphasizing its benefits, challenges, and future outlook in advancing sustainable practices. Furthermore, we underscore the critical need for continued research and development of advanced engineering techniques and genetic editing technologies tailored specifically for this bacterium.

| [1] |
Padda KP, Puri A, Zeng Q, et al. (2017) Effect of GFP-tagging on nitrogen fixation and plant growth promotion of an endophytic diazotrophic strain of Paenibacillus polymyxa. Botany 95: 933-942. https://doi.org/10.1139/cjb-2017-0056 
|
| [2] |
Ash C, Priest FG, Collins MD (1993) Molecular identification of rRNA group 3 bacilli (Ash, Farrow, Wallbanks and Collins) using a PCR probe test. Proposal for the creation of a new genus Paenibacillus. Antonie Van Leeuwenhoek 64: 253-260. https://doi.org/10.1007/BF00873085
|
| [3] |
Mahajan GB, Balachandran L (2017) Sources of antibiotics: Hot springs. Biochem Pharmacol 134: 35-41. https://doi.org/10.1016/j.bcp.2016.11.021
|
| [4] |
Tang Q, Puri A, Padda KP, et al. (2017) Biological nitrogen fixation and plant growth promotion of lodgepole pine by an endophytic diazotroph Paenibacillus polymyxa and its GFP-tagged derivative. Botany 95: 611-619. https://doi.org/10.1139/cjb-2016-0300
|
| [5] |
Padda KP, Puri A, Chanway CP (2017) Paenibacillus polymyxa: A prominent biofertilizer and biocontrol agent for sustainable agriculture. Agriculturally Important Microbes for Sustainable Agriculture . Singapore: Springer Singapore 165-191. https://doi.org/10.1007/978-981-10-5343-6_6
|
| [6] |
Mülner P, Schwarz E, Dietel K, et al. (2021) Fusaricidins, Polymyxins and volatiles produced by Paenibacillus polymyxa strains DSM 32871 and M1. Pathogens 10: 1485. https://doi.org/10.3390/pathogens10111485
|
| [7] |
Silva Dias BH, Jung SH, Castro Oliveira JVD, et al. (2021) C4 bacterial volatiles improve plant health. Pathogens 10: 682. https://doi.org/10.3390/pathogens10060682
|
| [8] |
Pandey AK, Barbetti MJ, Lamichhane JR (2023) Paenibacillus polymyxa. Trend Microbiol 31: 657-659. https://doi.org/10.1016/j.tim.2022.11.010
|
| [9] |
Kim SB, Timmusk S (2013) A simplified method for gene knockout and direct screening of recombinant clones for application in Paenibacillus polymyxa. Plos ONE 8: e68092. https://doi.org/10.1371/journal.pone.0068092
|
| [10] |
Meliawati M, Teckentrup C, Schmid J (2022) CRISPR-Cas9-mediated large cluster deletion and multiplex genome editing in Paenibacillus polymyxa. ACS Synth Biol 11: 77-84. https://doi.org/10.1021/acssynbio.1c00565
|
| [11] |
Zhang J, Zhao J, Fu Q, et al. (2024) Metabolic engineering of Paenibacillus polymyxa for effective production of 2,3-butanediol from poplar hydrolysate. Bioresour Technol 392: 130002. https://doi.org/10.1016/j.biortech.2023.130002
|
| [12] |
Ravagnan G, Meliawati M, Schmid J (2024) CRISPR-Cas9-Mediated genome editing in Paenibacillus polymyxa. Synthetic Biology: Methods and Protocols . New York, NY: Springer US 267-280. https://doi.org/10.1007/978-1-0716-3658-9_16
|
| [13] |
Wallner A, Antonielli L, Mesguida O, et al. (2024) Genomic diversity in Paenibacillus polymyxa: unveiling distinct species groups and functional variability. BMC Genomics 25: 720. https://doi.org/10.1186/s12864-024-10610-w
|
| [14] |
Maggi F, Giuliodori AM, Brandi A, et al. (2024) Pangenome analysis of Paenibacillus polymyxa strains reveals the existence of multiple and functionally distinct Paenibacillus species. Appl Environ Microbiol 90: e01740-24. https://doi.org/10.1128/aem.01740-24
|
| [15] |
Weselowski B, Nathoo N, Eastman AW, et al. (2016) Isolation, identification and characterization of Paenibacillus polymyxa CR1 with potentials for biopesticide, biofertilization, biomass degradation and biofuel production. BMC Microbiol 16: 244. https://doi.org/10.1186/s12866-016-0860-y
|
| [16] |
Langendries S, Goormachtig S (2021) Paenibacillus polymyxa, a Jack of all trades. Environ Microbiol 23: 5659-5669. https://doi.org/10.1111/1462-2920.15450
|
| [17] | Jeong H, Choi SK, Ryu CM, et al. (2019) Chronicle of a soil bacterium: Paenibacillus polymyxa E681 as a tiny guardian of plant and human health. Front Microbiol . https://doi.org/10.3389/fmicb.2019.00467 |
| [18] |
Cheng W, Yang J, Nie Q, et al. (2017) Volatile organic compounds from Paenibacillus polymyxa KM2501-1 control Meloidogyne incognita by multiple strategies. Sci Rep 7: 16213. https://doi.org/10.1038/s41598-017-16631-8
|
| [19] |
Rybakova D, Cernava T, Köberl M, et al. (2016) Endophytes-assisted biocontrol: novel insights in ecology and the mode of action of Paenibacillus. Plant Soil 405: 125-140. https://doi.org/10.1007/s11104-015-2526-1
|
| [20] |
Yegorenkova IV, Tregubova KV, Krasov AI, et al. (2021) Effect of exopolysaccharides of Paenibacillus polymyxa rhizobacteria on physiological and morphological variables of wheat seedlings. J Microbiol 59: 729-735. https://doi.org/10.1007/s12275-021-0623-9
|
| [21] |
Yuan Y, Xu QM, Yu SC, et al. (2020) Control of the polymyxin analog ratio by domain swapping in the nonribosomal peptide synthetase of Paenibacillus polymyxa. J Ind Microbiol Biotechnol 47: 551-562. https://doi.org/10.1007/s10295-020-02275-7
|
| [22] |
Zhang F, Li XL, Zhu SJ, et al. (2018) Biocontrol potential of Paenibacillus polymyxa against Verticillium dahliae infecting cotton plants. Biol Control 127: 70-77. https://doi.org/10.1016/j.biocontrol.2018.08.021
|
| [23] |
Galea CA, Han M, Zhu Y, et al. (2017) Characterization of the Polymyxin D Synthetase biosynthetic cluster and product profile of Paenibacillus polymyxa ATCC 10401. J Nat Prod 80: 1264-1274. https://doi.org/10.1021/acs.jnatprod.6b00807
|
| [24] |
Yegorenkova IV, Fomina AA, Tregubova KV, et al. (2018) Immunomodulatory activity of exopolysaccharide from the rhizobacterium Paenibacillus polymyxa CCM 1465. Arch Microbiol 200: 1471-1480. https://doi.org/10.1007/s00203-018-1564-5
|
| [25] |
Zhou L, Zhang T, Tang S, et al. (2020) Pan-genome analysis of Paenibacillus polymyxa strains reveals the mechanism of plant growth promotion and biocontrol. Antonie van Leeuwenhoek 113: 1539-1558. https://doi.org/10.1007/s10482-020-01461-y
|
| [26] |
Daud NS, Azam ZM, Othman NZ (2023) Optimization of medium compositions and functional characteristics of exopolysaccharide from Paenibacillus polymyxa ATCC 824. Biocatal Agric Biotechnol 49: 102656. https://doi.org/10.1016/j.bcab.2023.102656
|
| [27] |
Grady EN, MacDonald J, Liu L, et al. (2016) Current knowledge and perspectives of Paenibacillus: a review. Microb Cell Fact 15: 203. https://doi.org/10.1186/s12934-016-0603-7
|
| [28] |
Wu Y, Cai P, Jing X, et al. (2019) Soil biofilm formation enhances microbial community diversity and metabolic activity. Environ Int 132: 105116. https://doi.org/10.1016/j.envint.2019.105116
|
| [29] |
Zhou C, Guo J, Zhu L, et al. (2016) Paenibacillus polymyxa BFKC01 enhances plant iron absorption via improved root systems and activated iron acquisition mechanisms. Plant Physiol Biochem 105: 162-173. https://doi.org/10.1016/j.plaphy.2016.04.025
|
| [30] |
Daud NS, Mohd Din ARJ, Rosli MA, et al. (2019) Paenibacillus polymyxa bioactive compounds for agricultural and biotechnological applications. Biocatal Agric Biotechnol 18: 101092. https://doi.org/10.1016/j.bcab.2019.101092
|
| [31] |
Khan MS, Gao J, Chen X, et al. (2020) Isolation and characterization of plant growth-promoting endophytic bacteria Paenibacillus polymyxa SK1 from Lilium lancifolium. BioMed Res Int 2020: 8650957. https://doi.org/10.1155/2020/8650957
|
| [32] |
Mohd Din ARJ, Rosli MA, Mohamad Azam Z, et al. (2020) Paenibacillus polymyxa role involved in phosphate solubilization and growth promotion of Zea mays Under Abiotic Stress Condition. Proc Natl Acad Sci, India, Sect B Biol Sci 90: 63-71. https://doi.org/10.1007/s40011-019-01081-1
|
| [33] |
Li ZQ, Huang YL, Zhang J, et al. (2023) Ultrasound stimulated production of exopolysaccharide with anti-UV radiation activity by increasing cell permeability of Paenibacillus polymyxa. Process Biochem 126: 252-259. https://doi.org/10.1016/j.procbio.2023.01.010
|
| [34] |
Nasran HS, Mohd Yusof H, Halim M, et al. (2020) Optimization of protective agents for the freeze-drying of Paenibacillus polymyxa Kp10 as a potential biofungicide. Molecules 25: 2618. https://doi.org/10.3390/molecules25112618
|
| [35] |
Rani V, Bhatia A, Nain L, et al. (2021) Methane utilizing plant growth-promoting microbial diversity analysis of flooded paddy ecosystem of India. World J Microbiol Biotechnol 37: 56. https://doi.org/10.1007/s11274-021-03018-1
|
| [36] | Dal'Rio I, Lopes E dos S, Santaren KCF, et al. (2024) Co-inoculation of the endophytes Bacillus thuringiensis CAPE95 and Paenibacillus polymyxa CAPE238 promotes Tropaeolum majus L. growth and enhances its root bacterial diversity. Front Microbiol . https://doi.org/10.3389/fmicb.2024.1356891 |
| [37] |
Dobrzyński J, Naziębło A (2024) Paenibacillus as a biocontrol agent for fungal phytopathogens: is P. polymyxa the only one worth attention?. Microb Ecol 87: 134. https://doi.org/10.1007/s00248-024-02450-8
|
| [38] |
Ashrafian B, Hosseini-Abari A (2022) Investigation of bioactivity of unsaturated oligo‑galacturonic acids produced from apple waste by Alcaligenes faecalis AGS3 and Paenibacillus polymyxa S4 Pectinases. Sci Rep 12: 15830. https://doi.org/10.1038/s41598-022-20011-2
|
| [39] |
Li K, Jiang C, Tan H, et al. (2021) Identification and characterization of a novel glucomannanase from Paenibacillus polymyxa. 3 Biotech 11: 129. https://doi.org/10.1007/s13205-021-02676-0
|
| [40] |
Liu Q, Luo L, Zheng L (2018) Lignins: Biosynthesis and Biological Functions in Plants. Int J Mol Sci 19: 335. https://doi.org/10.3390/ijms19020335
|
| [41] |
Zhang W, Wang W, Wang J, et al. (2021) Isolation and characterization of a novel laccase for lignin degradation, LacZ1. Appl Environ Microbiol 87: e01355-21. https://doi.org/10.1128/AEM.01355-21
|
| [42] | Edith Ayala-Rodríguez A, Valdés-Rodríguez S, Enrique Olalde-Mathieu V, et al. (2024) Extracellular ligninases production and lignin degradation by Paenibacillus polymyxa. J Gen Appl Microbiol . https://doi.org/10.2323/jgam.2023.12.001 |
| [43] |
Parker GD, Plymale A, Hager J, et al. (2024) Studying microbially induced corrosion on glass using ToF-SIMS. Biointerphases 19: 051004. https://doi.org/10.1116/6.0003883
|
| [44] |
Tinôco D, Borschiver S, Coutinho PL, et al. (2021) Technological development of the bio-based 2,3-butanediol process. Biofuels Bioprod Bioref 15: 357-376. https://doi.org/10.1002/bbb.2173
|
| [45] | Sabra W, Quitmann H, Zeng AP, et al. (2011) 3.09-Microbial production of 2,3-Butanediol. Comprehensive Biotechnology . Burlington: Academic Press 87-97. https://doi.org/10.1016/B978-0-08-088504-9.00161-6 |
| [46] |
Joshi J, Langwald SV, Kruse O, et al. (2025) Immobilization of Paenibacillus polymyxa with biopolymers to enhance the production of 2,3-butanediol. Microb Cell Fact 24: 15. https://doi.org/10.1186/s12934-024-02633-5
|
| [47] |
Tinôco D, De Castro AM, Seldin L, et al. (2021) Production of (2R,3R)-butanediol by Paenibacillus polymyxa PM 3605 from crude glycerol supplemented with sugarcane molasses. Process Biochem 106: 88-95. https://doi.org/10.1016/j.procbio.2021.03.030
|
| [48] |
Zhang L, Chen S, Xie H, et al. (2012) Efficient acetoin production by optimization of medium components and oxygen supply control using a newly isolated Paenibacillus polymyxa CS107. J of Chemical Tech Biotech 87: 1551-1557. https://doi.org/10.1002/jctb.3791
|
| [49] |
Tinôco D, Pateraki C, Koutinas AA, et al. (2021) Bioprocess development for 2,3-Butanediol production by Paenibacillus strains. ChemBioEng Rev 8: 44-62. https://doi.org/10.1002/cben.202000022
|
| [50] | Stoklosa RJ, Latona RJ, Johnston DB (2022) Assessing oxygen limiting fermentation conditions for 2,3-butanediol production from Paenibacillus polymyxa. Front Chem Eng . https://doi.org/10.3389/fceng.2022.1038311 |
| [51] |
Tinôco D, Freire DMG (2023) Scale-up of 2,3-butanediol production by Paenibacillus peoriae NRRL BD-62 using constant oxygen transfer rate-based strategy. Fuel 340: 127603. https://doi.org/10.1016/j.fuel.2023.127603
|
| [52] |
Chen Q, Song Y, An Y, et al. (2024) Mechanisms and impact of rhizosphere microbial metabolites on crop health, traits, functional components: A comprehensive review. Molecules 29: 5922. https://doi.org/10.3390/molecules29245922
|
| [53] |
Cho G, Kim DR, Jeon CW, et al. (2020) Draft genome sequence data of Paenibacillus Polymyxa strain TH2H2, isolated from a tomato flower in Korea. Data Brief 31: 105824. https://doi.org/10.1016/j.dib.2020.105824
|
| [54] |
Chen N, Cai P, Zhang D, et al. (2024) Metabolic engineering of “last-line antibiotic” colistin in Paenibacillus polymyxa. Metab Eng 85: 35-45. https://doi.org/10.1016/j.ymben.2024.07.005
|
| [55] |
Li H, E WH, Zhao D, et al. (2024) Response of Paenibacillus polymyxa SC2 to the stress of polymyxin B and a key ABC transporter YwjA involved. Appl Microbiol Biotechnol 108: 17. https://doi.org/10.1007/s00253-023-12916-3
|
| [56] |
Singh B, Kumar N, Yadav A, et al. (2025) Harnessing the power of bacteriocins: A comprehensive review on sources, mechanisms, and applications in food preservation and safety. Curr Microbiol 82: 174. https://doi.org/10.1007/s00284-025-04155-8
|
| [57] |
El-Sharoud WM, Zalma SA, Yousef AE (2022) Inducing the production of the bacteriocin paenibacillin by Paenibacillus polymyxa through application of environmental stresses with relevance to milk bio-preservation. Int J Food Microbiol 371: 109637. https://doi.org/10.1016/j.ijfoodmicro.2022.109637
|
| [58] |
Cunha I de CM da, Silva AVR da, Boleta EHM, et al. (2024) The interplay between the inoculation of plant growth-promoting rhizobacteria and the rhizosphere microbiome and their impact on plant phenotype. Microbiol Res 283: 127706. https://doi.org/10.1016/j.micres.2024.127706
|
| [59] |
Kumar S, Ujor VC (2022) Complete genome sequence of Paenibacillus polymyxa DSM 365, a soil bacterium of agricultural and industrial importance. Microbiol Resour Announce 11: e00329-22. https://doi.org/10.1128/mra.00329-22
|
| [60] |
Zalila-Kolsi I, Ben Mahmoud A, Ali H, et al. (2016) Antagonist effects of Bacillus spp. strains against Fusarium graminearum for protection of durum wheat (Triticum turgidum L. subsp. durum). Microbio Res 192: 148-158. https://doi.org/10.1016/j.micres.2016.06.012
|
| [61] |
Addesso R, Sofo A, Amato M (2023) Rhizosheath: roles, formation processes and investigation methods. Soil Systems 7: 106. https://doi.org/10.3390/soilsystems7040106
|
| [62] |
Xu F, Liao H, Yang J, et al. (2023) Auxin-producing bacteria promote barley rhizosheath formation. Nat Commun 14: 5800. https://doi.org/10.1038/s41467-023-40916-4
|
| [63] |
Li X, Ma S, Meng Y, et al. (2023) Characterization of antagonistic bacteria Paenibacillus polymyxa ZYPP18 and the effects on plant growth. Plants 12: 2504. https://doi.org/10.3390/plants12132504
|
| [64] |
Wang H, Wang N, Tan Y, et al. (2023) YLC1: a promising antagonistic strain for biocontrol of pv., causing kiwifruit bacterial canker. Pest Manage Sci 79: 4357-4366. https://doi.org/10.1002/ps.7633
|
| [65] |
Shi Q, Zhang J, Fu Q, et al. (2024) Biocontrol efficacy and induced resistance of Paenibacillus polymyxa J2-4 against Meloidogyne incognita infection in Cucumber. Phytopathology® 114: 538-548. https://doi.org/10.1094/PHYTO-03-23-0091-R
|
| [66] | Abdelkhalek A, Al-Askar AA, Elbeaino T, et al. (2022) Protective and curative activities of Paenibacillus polymyxa against Zucchini yellow mosaic virus infestation in Squash Plants. Biology (Basel) 11: 1150. https://doi.org/10.3390/biology11081150 |
| [67] | Abdel Latef AAH, Zaid A, Abo-Baker A-BA-E, et al. (2020) Mitigation of copper stress in maize by inoculation with Paenibacillus polymyxa and Bacillus circulans. Plants (Basel) 9: 1513. https://doi.org/10.3390/plants9111513 |
| [68] |
Wang C, Pei J, Li H, et al. (2024) Mechanisms on salt tolerant of Paenibacillus polymyxa SC2 and its growth-promoting effects on maize seedlings under saline conditions. Microbiol Res 282: 127639. https://doi.org/10.1016/j.micres.2024.127639
|
| [69] |
Jozay M, Zarei H, Khorasaninejad S, et al. (2024) Exploring the impact of plant growth-promoting bacteria in alleviating stress on Aptenia cordifolia subjected to irrigation with recycled water in multifunctional external green walls. BMC Plant Biol 24: 802. https://doi.org/10.1186/s12870-024-05511-9
|
| [70] |
He Z, Kisla D, Zhang L, et al. (2007) Isolation and identification of a Paenibacillus polymyxa strain that coproduces a novel lantibiotic and polymyxin. Appl Environ Microbiol 73: 168-178. https://doi.org/10.1128/AEM.02023-06
|
| [71] |
Chiu S, Hancock AM, Schofner BW, et al. (2022) Causes of polymyxin treatment failure and new derivatives to fill the gap. J Antibiot (Tokyo) 75: 593-609. https://doi.org/10.1038/s41429-022-00561-3
|
| [72] |
Hagh Ranjbar H, Hosseini-Abari A, Ghasemi SM, et al. (2023) Antibacterial activity of epsilon-poly-l-lysine produced by Stenotrophomonas maltophilia HS4 and Paenibacillus polymyxa HS5, alone and in combination with bacteriophages. Microbiology (Reading) 169: 001363. https://doi.org/10.1099/mic.0.001363
|
| [73] |
Mokhtar NFK, Hashim AM, Hanish I, et al. (2020) The discovery of new antilisterial proteins from Paenibacillus polymyxa Kp10 via genome mining and mass spectrometry. Front Microbiol 11: 960. https://doi.org/10.3389/fmicb.2020.00960
|
| [74] |
Thapa RK, Kim JO, Kim J (2023) Antimicrobial strategies for topical biofilm-based wound infections: past, present, and future. J Pharm Investig 53: 627-641. https://doi.org/10.1007/s40005-023-00628-9
|
| [75] |
Ghoreishi FS, Roghanian R, Emtiazi G (2022) Novel chronic wound healing by anti-biofilm peptides and protease. Adv Pharm Bull 12: 424-436. https://doi.org/10.34172/apb.2022.047
|
| [76] | Zhang X, Chen G, Yu Y, et al. (2020) Bioinspired adhesive and antibacterial microneedles for versatile transdermal drug delivery. Research (Wash D C) 2020: 3672120. https://doi.org/10.34133/2020/3672120 |
| [77] |
Xie WY, Shen HL, Yan ZM, et al. (2024) Paenibacillus exopolysaccharide alleviates Malassezia-induced skin damage: Enhancing skin barrier function, regulating immune responses, and modulating microbiota. Int J Biol Macromol 278: 135404. https://doi.org/10.1016/j.ijbiomac.2024.135404
|
| [78] |
Yasuzawa T, Nishi R, Ishitani S, et al. (2022) Effects of enzamin, a microbial product, on alterations of intestinal microbiota induced by a high-fat diet. Nutrients 14: 4743. https://doi.org/10.3390/nu14224743
|
| [79] |
Rütering M, Cress BF, Schilling M, et al. (2017) Tailor-made exopolysaccharides—CRISPR-Cas9 mediated genome editing in Paenibacillus polymyxa. Synth Biol 2: ysx007. https://doi.org/10.1093/synbio/ysx007
|
| [80] |
Okonkwo CC, Ujor V, Cornish K, et al. (2020) Inactivation of the levansucrase gene in Paenibacillus polymyxa DSM 365 diminishes exopolysaccharide biosynthesis during 2,3-butanediol fermentation. Appl Environ Microbiol 86: e00196-20. https://doi.org/10.1128/AEM.00196-20
|
| [81] | Irla M, Heggeset TMB, Nærdal I, et al. (2016) Genome-based genetic tool development for Bacillus methanolicus: Theta- and rolling circle-replicating plasmids for inducible gene expression and application to methanol-based cadaverine production. Front Microbiol . https://doi.org/10.3389/fmicb.2016.01481 |
| [82] |
Chen L, Duan L, Sun M, et al. (2023) Current trends and insights on EMS mutagenesis application to studies on plant abiotic stress tolerance and development. Front Plant Sci 13: 1052569. https://doi.org/10.3389/fpls.2022.1052569
|
| [83] |
Soliman EAM, A.H. Aly N, Abosereh NA, et al. (2020) Mutagenic improvement of some bacillus strains for enhanced alpha amylase production. Int J Curr Microbiol App Sci 9: 1266-1277. https://doi.org/10.20546/ijcmas.2020.909.155
|
| [84] |
Hoyos-Manchado R, Villa-Consuegra S, Berraquero M, et al. (2020) Mutational Analysis of N-Ethyl-N-Nitrosourea (ENU) in the Fission Yeast Schizosaccharomyces pombe. G3 Genes Genomes Genet 10: 917-923. https://doi.org/10.1534/g3.119.400936
|
| [85] |
Lawley PD, Thatcher CJ (1970) Methylation of deoxyribonucleic acid in cultured mammalian cells by N-methyl-N′-nitro-N-nitrosoguanidine. The influence of cellular thiol concentrations on the extent of methylation and the 6-oxygen atom of guanine as a site of methylation. Biochem J 116: 693-707. https://doi.org/10.1042/bj1160693
|
| [86] |
Liu H, Wang J, Sun H, et al. (2020) Transcriptome profiles reveal the growth-promoting mechanisms of Paenibacillus polymyxa YC0136 on Tobacco (Nicotiana tabacum L.). Front Microbiol 11: 584174. https://doi.org/10.3389/fmicb.2020.584174
|
| [87] |
Timmusk S, Pall T, Raz S, et al. (2023) The potential for plant growth-promoting bacteria to impact crop productivity in future agricultural systems is linked to understanding the principles of microbial ecology. Front Microbiol 14: 1141862. https://doi.org/10.3389/fmicb.2023.1141862
|
| [88] |
Ryu CM, Kim J, Choi O, et al. (2006) Improvement of biological control capacity of Paenibacillus polymyxa E681 by seed pelleting on sesame. Biol Control 39: 282-289. https://doi.org/10.1016/j.biocontrol.2006.04.014
|
| [89] |
Wang Z, Wang S, Yang H (2024) Understanding the pathogenesis, biocontrol mechanisms, and factors influencing biocontrol effectiveness for soil-borne diseases in panax plants. Microorganisms 12: 2278. https://doi.org/10.3390/microorganisms12112278
|
| [90] |
Yuan P, Chen Z, Xu M, et al. (2024) Microbial cell factories using Paenibacillus : status and perspectives. Critical Rev Biotechnol 44: 1386-1402. https://doi.org/10.1080/07388551.2023.2289342
|
| [91] |
Kumar P, Kumar V, Kumar R, et al. (2020) Fabrication and characterization of ceftizoxime-loaded pectin nanocarriers. Nanomaterials 10: 1452. https://doi.org/10.3390/nano10081452
|
| [92] |
Zhang Q, Xing C, Li S, et al. (2021) In vitro antagonism and biocontrol effects of Paenibacillus polymyxa JY1-5 against Botrytis cinerea in tomato. Biol Control 160: 104689. https://doi.org/10.1016/j.biocontrol.2021.104689
|
| [93] |
Choi SK, Park SY, Kim R, et al. (2009) Identification of a polymyxin synthetase gene cluster of Paenibacillus polymyxa and heterologous expression of the gene in Bacillus subtilis. J Bacteriol 191: 3350-3358. https://doi.org/10.1128/JB.01728-08
|
| [94] |
Živič Z, Lipoglavšek L, Lah J, et al. (2025) A single vector system for tunable and homogeneous dual gene expression in Escherichia coli. Sci Rep 15: 99. https://doi.org/10.1038/s41598-024-83628-5
|
| [95] |
Rappleye CA (2023) Targeted gene deletions in the dimorphic fungal pathogen Histoplasma using an optimized episomal CRISPR/Cas9 system. mSphere 8: e00178-23. https://doi.org/10.1128/msphere.00178-23
|
| [96] | Kim MS, Kim HR, Jeong DE, et al. (2021) Cytosine base editor-mediated multiplex genome editing to accelerate discovery of novel antibiotics in Bacillus subtilis and Paenibacillus polymyxa. Front Microbiol . https://doi.org/10.3389/fmicb.2021.691839 |
| [97] |
Yan D, Tajima H, Cline LC, et al. (2022) Genetic modification of flavone biosynthesis in rice enhances biofilm formation of soil diazotrophic bacteria and biological nitrogen fixation. Plant Biotechnol J 20: 2135-2148. https://doi.org/10.1111/pbi.13894
|
| [98] |
Kumar SS, Sabu A (2019) Fibrinolytic enzymes for thrombolytic therapy. Therapeutic Enzymes: Function and Clinical Implications . Singapore: Springer Singapore 345-381. https://doi.org/10.1007/978-981-13-7709-9_15
|
| [99] |
Schilling C, Koffas MAG, Sieber V, et al. (2020) Novel prokaryotic CRISPR-Cas12a-Based tool for programmable transcriptional activation and repression. ACS Synth Biol 9: 3353-3363. https://doi.org/10.1021/acssynbio.0c00424
|
| [100] |
Li Y, Chen S (2019) Fusaricidin produced by Paenibacillus polymyxa WLY78 Induces systemic resistance against Fusarium Wilt of Cucumber. IJMS 20: 5240. https://doi.org/10.3390/ijms20205240
|
| [101] |
He X, Li Q, Wang N, et al. (2021) Effects of an EPS biosynthesis gene cluster of Paenibacillus polymyxa WLY78 on biofilm formation and nitrogen fixation under aerobic conditions. Microorganisms 9: 289. https://doi.org/10.3390/microorganisms9020289
|
| [102] |
Liu H, Li Y, Ge K, et al. (2021) Interactional mechanisms of Paenibacillus polymyxa SC2 and pepper (Capsicum annuum L.) suggested by transcriptomics. BMC Microbiol 21: 70. https://doi.org/10.1186/s12866-021-02132-2
|
| [103] |
Ju JH, Jo MH, Heo SY, et al. (2023) Production of highly pure R,R-2,3-butanediol for biological plant growth promoting agent using carbon feeding control of Paenibacillus polymyxa MDBDO. Microb Cell Fact 22: 121. https://doi.org/10.1186/s12934-023-02133-y
|
| [104] |
Chen Z, Shen M, Mao C, et al. (2021) A type I restriction modification system influences genomic evolution driven by horizontal gene transfer in Paenibacillus polymyxa. Front Microbiol 12: 709571. https://doi.org/10.3389/fmicb.2021.709571
|
| [105] | Engwa G Genetic engineering on microorganism: the ecological and bioethical implications (2014)1: 27-33. |
| [106] |
Dawkins K, Esiobu N (2018) The invasive brazilian pepper tree (Schinus terebinthifolius) is colonized by a root microbiome enriched with alphaproteobacteria and unclassified spartobacteria. Front Microbiol 9: 876. https://doi.org/10.3389/fmicb.2018.00876
|
| [107] |
O'Sullivan GM, Philips JG, Mitchell HJ, et al. (2022) 20 years of legislation - how australia has responded to the challenge of regulating genetically modified organisms in the clinic. Front Med 9: 883434. https://doi.org/10.3389/fmed.2022.883434
|
| [108] |
Nielsen J, Keasling JD (2016) Engineering cellular metabolism. Cell 164: 1185-1197. https://doi.org/10.1016/j.cell.2016.02.004
|
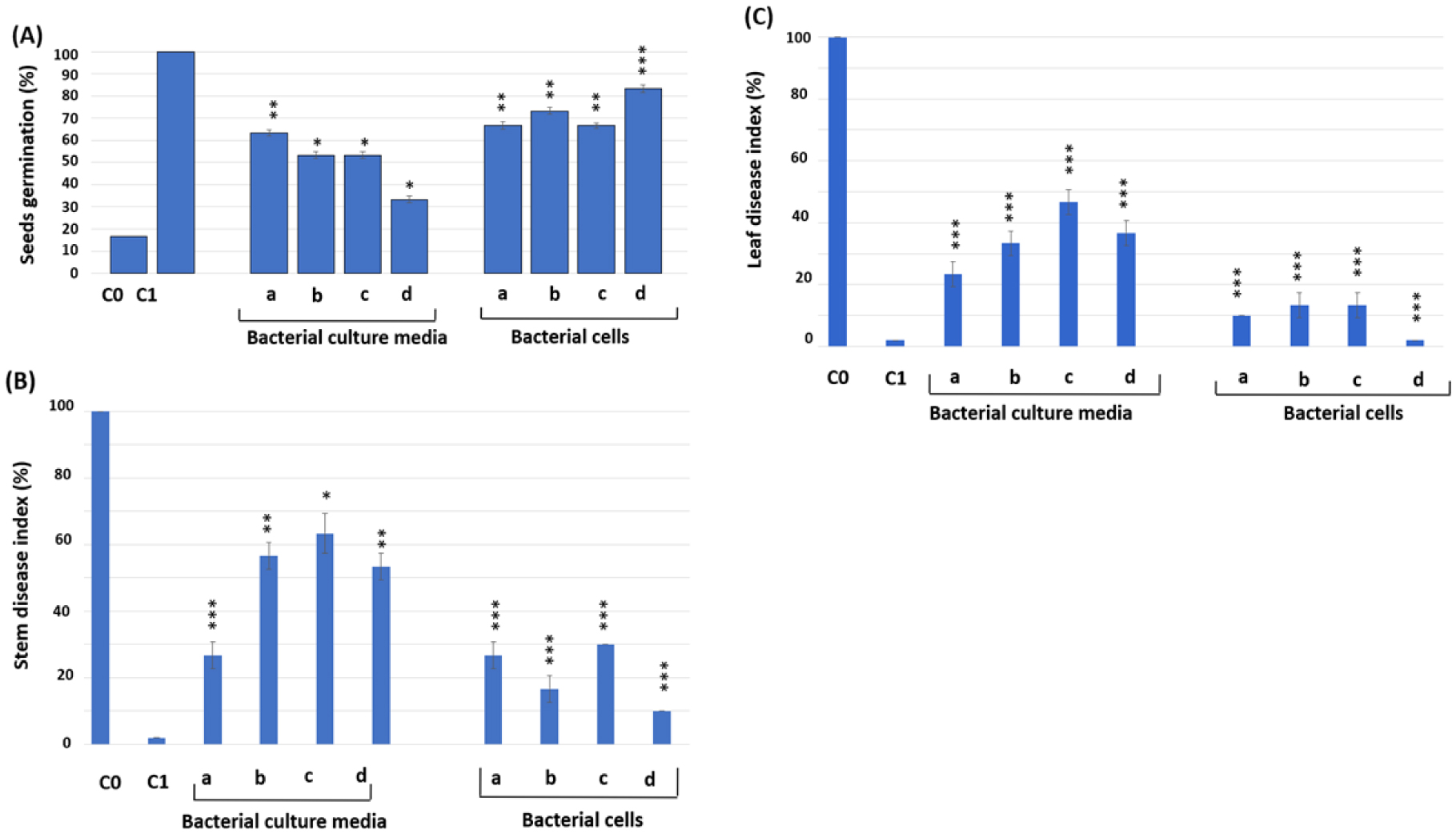
Figures(2) / Tables(3)

Imen Zalila-Kolsi, Ray Al-Barazie. Advancing sustainable practices with Paenibacillus polymyxa: From soil health to medical applications and molecular engineering[J]. AIMS Microbiology, 2025, 11(2): 338-368. doi: 10.3934/microbiol.2025016







 DownLoad:
DownLoad: