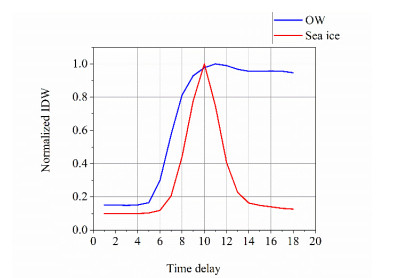
Global navigation satellite system reflectometry (GNSS-R) is based on satellite signals' multipath interference effect and has developed as one of the important remote sensing technologies in sea ice detection. An isometric mapping (ISOMAP)-based method is proposed in this paper as a development in sea ice detection approaches. The integral delay waveforms (IDWs), selected from February to April in 2018, derived from TechDemoSat-1 (TDS-1) Delay-Doppler maps (DDMs) are applied to open water and sea ice classification. In the first, the model for extracting low-dimensional coordinates of IDWs employs the randomly selected 187666 IDW samples, which are 30% of the whole IDW dataset. Then, low-dimensional coordinates of IDWs are used to train three different classifiers of support vector machine (SVM) and gradient boosting decision tree (GBDT), linear discriminant algorithm (LDA) and K-nearest neighbors (KNN) for determining the sea ice and sea water. The remaining 437889 samples, about 70% of the whole datasets, are used to validate with the ground surface type from the National Snow and Ice Data Center (NSIDC) data provided by the National Oceanic and Atmospheric Administration (NOAA). The algorithm performance is evaluated, and the overall accuracy of SVM, GBDT, LDA and KNN are 99.44%, 85.58%, 91.88% and 98.82%, respectively. The sea ice detection methods are developed, and the accuracy of detection is improved in this paper.
Citation: Yuan Hu, Zhihao Jiang, Xintai Yuan, Xifan Hua, Wei Liu. Isometric mapping algorithm based GNSS-R sea ice detection[J]. Metascience in Aerospace, 2024, 1(1): 38-52. doi: 10.3934/mina.2024002
Global navigation satellite system reflectometry (GNSS-R) is based on satellite signals' multipath interference effect and has developed as one of the important remote sensing technologies in sea ice detection. An isometric mapping (ISOMAP)-based method is proposed in this paper as a development in sea ice detection approaches. The integral delay waveforms (IDWs), selected from February to April in 2018, derived from TechDemoSat-1 (TDS-1) Delay-Doppler maps (DDMs) are applied to open water and sea ice classification. In the first, the model for extracting low-dimensional coordinates of IDWs employs the randomly selected 187666 IDW samples, which are 30% of the whole IDW dataset. Then, low-dimensional coordinates of IDWs are used to train three different classifiers of support vector machine (SVM) and gradient boosting decision tree (GBDT), linear discriminant algorithm (LDA) and K-nearest neighbors (KNN) for determining the sea ice and sea water. The remaining 437889 samples, about 70% of the whole datasets, are used to validate with the ground surface type from the National Snow and Ice Data Center (NSIDC) data provided by the National Oceanic and Atmospheric Administration (NOAA). The algorithm performance is evaluated, and the overall accuracy of SVM, GBDT, LDA and KNN are 99.44%, 85.58%, 91.88% and 98.82%, respectively. The sea ice detection methods are developed, and the accuracy of detection is improved in this paper.

| [1] |
Screen JA, Simmonds I (2010) The central role of diminishing sea ice in recent Arctic temperature amplification. Nature 464: 1334–1337. https://doi.org/10.1038/nature09051 doi: 10.1038/nature09051

|
| [2] | Park JW, Korosov AA, Babiker M, et al. (2019) Classification of sea ice types in Sentinel-1 SAR images. Cryosphere Discuss. 2019: 1–23. |
| [3] |
Dabboor M, Geldsetzer T (2014) Towards sea ice classification using simulated RADARSAT Constellation Mission compact polarimetric SAR imagery. Remote Sens Environ 140: 189–195. https://doi.org/10.1016/j.rse.2013.08.035 doi: 10.1016/j.rse.2013.08.035
|
| [4] |
Cardellach E, Fabra F, Nogués-Correig O, et al. (2011) GNSS-R ground-based and airborne campaigns for ocean, land, ice, and snow techniques: Application to the GOLD-RTR data sets. Radio Sci 46: 1–16. https://doi.org/10.1029/2011rs004683 doi: 10.1029/2011rs004683
|
| [5] | Martin-Neira M (1993) A passive reflectometry and interferometry system (PARIS): Application to ocean altimetry. ESA J 17: 331–355. |
| [6] | Hall C, Cordey R (1988) Multistatic scatterometry, International Geoscience and Remote Sensing Symposium, Remote Sensing: Moving Toward the 21st Century', IEEE. |
| [7] |
Liu Y, Collett I, Morton YJ (2019) Application of Neural Network to GNSS-R Wind Speed Retrieval. IEEE T Geosc Remote 57: 9756–9766. https://doi.org/10.1109/tgrs.2019.2929002 doi: 10.1109/tgrs.2019.2929002
|
| [8] |
Garrison JL, Komjathy A, Zavorotny VU, et al. (2022) Wind speed measurement using forward scattered GPS signals. IEEE T Geosc Remote 40: 50–65. https://doi.org/10.1109/36.981349 doi: 10.1109/36.981349
|
| [9] |
Clarizia M P, Ruf C S, Jales P, et al. (2014) Gommenginger. Spaceborne GNSS-R Minimum Variance Wind Speed Estimator. IEEE T Geosc Remote 52: 6829–6843. https://doi.org/10.1109/Tgrs.2014.2303831 doi: 10.1109/Tgrs.2014.2303831
|
| [10] |
Yu K, Li Y, Chang X (2019) Chang. Snow Depth Estimation Based on Combination of Pseudorange and Carrier Phase of GNSS Dual-Frequency Signals. IEEE T Geosc Remote 57: 1817–1828. https://doi.org/10.1109/tgrs.2018.2869284 doi: 10.1109/tgrs.2018.2869284
|
| [11] |
Camps A, Park H, Pablos M, et al. (2016) Sensitivity of GNSS-R Spaceborne Observations to Soil Moisture and Vegetation. IEEE J-Stars 9: 4730–4742. https://doi.org/10.1109/jstars.2016.2588467 doi: 10.1109/jstars.2016.2588467
|
| [12] |
Rodriguez-Alvarez N, Bosch-Lluis X, Camps A, et al. (2009) Soil Moisture Retrieval Using GNSS-R Techniques: Experimental Results Over a Bare Soil Field. IEEE T Geosc Remote 47: 3616–3624. https://doi.org/10.1109/tgrs.2009.2030672 doi: 10.1109/tgrs.2009.2030672
|
| [13] |
Di Simone A, Park H, Riccio D, et al. (2017) Sea Target Detection Using Spaceborne GNSS-R Delay-Doppler Maps: Theory and Experimental Proof of Concept Using TDS-1 Data. IEEE J-Stars 10: 4237–4255. https://doi.org/10.1109/jstars.2017.2705350 doi: 10.1109/jstars.2017.2705350
|
| [14] |
Alonso-Arroyo A, Zavorotny VU, Camps A (2017) Sea ice detection using UK TDS-1 GNSS-R data. IEEE J-Stars 55: 4989–5001. https://doi.org/10.1109/TGRS.2017.2699122 doi: 10.1109/TGRS.2017.2699122
|
| [15] |
Yan Q, Huang W (2016) Sea ice detection from GNSS-R delay-Doppler map. 2016 17th International Symposium on Antenna Technology and Applied Electromagnetics (ANTEM), 2016: 1–2. https://doi.org/10.1109/ANTEM.2016.7550123 doi: 10.1109/ANTEM.2016.7550123
|
| [16] |
Yan Q, Huang W (2016) Spaceborne GNSS-R Sea Ice Detection Using Delay-Doppler Maps: First Results From the U.K. TechDemoSat-1 Mission. IEEE J-Stars 9: 4795–4801. https://doi.org/10.1109/jstars.2016.2582690 doi: 10.1109/jstars.2016.2582690
|
| [17] |
Munoz-Martin J F, Perez A, Camps A, et al. (2020) Snow and Ice Thickness Retrievals Using GNSS-R: Preliminary Results of the MOSAiC Experiment. Rem Sen 12: 4038. https://doi.org/10.3390/rs12244038 doi: 10.3390/rs12244038
|
| [18] |
Zavorotny VU, Voronovich AG (2000) Scattering of GPS signals from the ocean with wind remote sensing application. IEEE T Geosci Remote 38: 951–964. https://doi.org/10.1109/36.841977 doi: 10.1109/36.841977
|
| [19] |
Zhu Y, Yu K, Zou J, et al. (2017) Sea Ice Detection Based on Differential Delay-Doppler Maps from UK TechDemoSat-1. Sensors (Basel) 17: 1614. https://doi.org/10.3390/s17071614 doi: 10.3390/s17071614
|
| [20] |
Yan Q, Huang W (2018) Sea Ice Sensing From GNSS-R Data Using Convolutional Neural Networks. IEEE Geosci Remote Sens Lett 15: 1510–1514. https://doi.org/10.1109/lgrs.2018.2852143 doi: 10.1109/lgrs.2018.2852143
|
| [21] |
Hu Y, Jiang Z, Liu W, et al. (2023) GNSS-R Sea Ice Detection Based on Linear Discriminant Analysis. IEEE T Geosci Remote. https://doi.org/10.1109/TGRS.2023.3269088 doi: 10.1109/TGRS.2023.3269088
|
| [22] | Eustice D, Baylis C, Marks RJ (2015) Woodward's ambiguity function: From foundations to applications, 2015 Texas Symposium on Wireless and Microwave Circuits and Systems (WMCS); 2015: IEEE. 2015: 1–17. https://doi.org/10.1109/WMCaS.2015.7233208 |
| [23] | Jales P, Unwin M (2015) MERRByS product manual: GNSS reflectometry on TDS-1 with the SGR-ReSI. Surrey Satellite Technol. Ld. Guildford, UK. |
| [24] |
Samko O, Marshall A D, Rosin P L (2006) Rosin. Selection of the optimal parameter value for the Isomap algorithm. Pattern Recogn Lett 27: 968–979. https://doi.org/10.1016/j.patrec.2005.11.017 doi: 10.1016/j.patrec.2005.11.017
|
| [25] |
Heinzel S, Berg D, Gasser T, et al. (2019) Update of the MDS research criteria for prodromal Parkinson's disease. Mov Disord. 34: 1464–1470. https://doi.org/10.1002/mds.27802 doi: 10.1002/mds.27802
|
| [26] |
Hougardy S (2010) The Floyd–Warshall algorithm on graphs with negative cycles. Inform Process Lett 110: 279–281. https://doi.org/10.1016/j.ipl.2010.02.001 doi: 10.1016/j.ipl.2010.02.001
|
| [27] |
Cortes C, Vapnik V (1995) Support-Vector Networks. Mach Learn 20: 273–297. https://doi.org/10.1007/Bf00994018 doi: 10.1007/Bf00994018
|
| [28] |
Liang W, Luo S, Zhao G, et al. (2020) Predicting Hard Rock Pillar Stability Using GBDT, XGBoost, and LightGBM Algorithms. Mathematics 8: 765. https://doi.org/10.3390/math8050765 doi: 10.3390/math8050765
|
| [29] | Izenman A (2013) Linear Discriminant Analysis, In: Modern Multivariate Statistical Techniques, Springer Texts in Statistics: Springer, 237–280. |
| [30] |
Turk M, Pentland A (1991) Eigenfaces for recognition. J Cogn Neurosci 3: 71–86. https://doi.org/10.1162/jocn.1991.3.1.71 doi: 10.1162/jocn.1991.3.1.71
|
| [31] |
Wang L, Zhang Y, Feng J (2005) On the Euclidean distance of images. IEEE Trans Pattern Anal Mach Intel 27: 1334–1339. https://doi.org/10.1109/TPAMI.2005.165 doi: 10.1109/TPAMI.2005.165
|
| [32] | Guo G, Wang H, Bell D, et al. (2003) KNN model-based approach in classification, On The Move to Meaningful Internet Systems 2003: CoopIS, DOA, and ODBASE: OTM Confederated International Conferences, CoopIS, DOA, and ODBASE 2003, Catania, Sicily, Italy, November 3–7. Proceedings; Springer. |
| [33] | Kraemer HC (2014) Kappa coefficient. Wiley StatsRef: statistics reference online: 1–4. https://doi.org/10.1002/9781118445112.stat00365.pub2 |
| [34] |
Alonso-Arroyo A, Zavorotny V U, Camps A (2017) Sea Ice Detection Using U.K. TDS-1 GNSS-R Data. IEEE T Geosci Remot 55: 4989–5001. https://doi.org/10.1109/tgrs.2017.2699122 doi: 10.1109/tgrs.2017.2699122
|
| [35] |
Yan Q, Huang W (2019) Detecting Sea Ice From TechDemoSat-1 Data Using Support Vector Machines With Feature Selection. IEEE J--STARS 12: 1409–1416. https://doi.org/10.1109/jstars.2019.2907008 doi: 10.1109/jstars.2019.2907008
|
| [36] |
Yan Q, Huang W (2017) Neural Networks Based Sea Ice Detection and Concentration Retrieval From GNSS-R Delay-Doppler Maps. IEEE J--STARS 10: 3789–3798. https://doi.org/10.1109/jstars.2017.2689009 doi: 10.1109/jstars.2017.2689009
|





Figures(6) / Tables(2)
Yuan Hu, Zhihao Jiang, Xintai Yuan, Xifan Hua, Wei Liu. Isometric mapping algorithm based GNSS-R sea ice detection[J]. Metascience in Aerospace, 2024, 1(1): 38-52. doi: 10.3934/mina.2024002







 DownLoad:
DownLoad: