Citation: Tika Ram Lamichhane, Hari Prasad Lamichhane. Heat conduction by thyroid hormone receptors[J]. AIMS Biophysics, 2018, 5(4): 245-256. doi: 10.3934/biophy.2018.4.245

| [1] |
Brent GA (2012) Mechanisms of thyroid hormone action. J Clin Invest 122: 3035–3043. doi: 10.1172/JCI60047

|
| [2] |
Souza PCT, Puhl AC, Martínez L, et al. (2014) Identification of a new hormone-binding site on the surface of thyroid hormone receptor. Mol Endocrinol 28: 534–545. doi: 10.1210/me.2013-1359
|
| [3] |
Bochukova E, Schoenmakers N, Agostini M, et al. (2012) A mutation in the thyroid hormone receptor alpha gene. New Eng J Med 366: 243–249. doi: 10.1056/NEJMoa1110296
|
| [4] |
Brent GA (1994) The molecular basis of thyroid hormone action. Eng J Med 331: 847–853. doi: 10.1056/NEJM199409293311306
|
| [5] | Leitner DM (2008) Energy flow in proteins. Annu Rev Phys Chem 59: 233–259. |
| [6] |
Prabhu NV, Sharp KA (2005) Heat capacity in proteins. Annu Rev Phys Chem 56: 521–548. doi: 10.1146/annurev.physchem.56.092503.141202
|
| [7] |
Privalov PL, Tiktopulo EI, Venyaminov SY, et al. (1989) Heat capacity and conformation of proteins in the denatured state. J Mol Biol 205: 737–750. doi: 10.1016/0022-2836(89)90318-5
|
| [8] |
Yu X, Leitner DM (2005) Heat flow in proteins: Computation of thermal transport coefficients. J Chem Phys 122: 054902. doi: 10.1063/1.1830431
|
| [9] |
Lervik A, Bresme F, Kjelstrup S, et al. (2010) Heat transfer in protein-water interfaces. Phys Chem Chem Phys 12: 1610–1617. doi: 10.1039/b918607g
|
| [10] | Helbing J, Devereux M, Nienhaus K, et al. (2011) Temperature dependence of the heat diffusivity of proteins. J Phys Chem A 116: 2620–2628. |
| [11] |
Nosé S (1984) A molecular dynamics method for simulations in the canonical ensemble. Mol Phys 52: 255–268. doi: 10.1080/00268978400101201
|
| [12] | Suurkuusk J (1974) Specific heat measurements on lysozyme, chymotrypsinogen, and ovalbumin in aqueous solution and in solid state. Acta Chem Scand B 28: 409–417. |
| [13] | Yang PH, Rupley JA (1979) Protein-water interactions. Heat capacity of the lysozyme-water system. Biochemistry 18: 2654–2661. |
| [14] |
Lervik A, Bresme F, Kjelstrup S (2009) Heat transfer in soft nanoscale interfaces: The influence of interface curvature. Soft Matter 5: 2407–2414. doi: 10.1039/b817666c
|
| [15] |
Nascimento AS, Dia SMG, Nunes FM, et al. (2006) Structural rearrangements in the thyroid hormone receptor hinge domain and their putative role in the receptor function. J Mol Biol 360: 586–598. doi: 10.1016/j.jmb.2006.05.008
|
| [16] |
Sandler B, Webb P, Apriletti JW, et al. (2004) Thyroxine-thyroid hormone receptor interactions. J Biol Chem 279: 55801–55808. doi: 10.1074/jbc.M410124200
|
| [17] |
Souza PCT, Puhl AC, Martínez L, et al. (2014) Identification of a new hormone-binding site on the surface of thyroid hormone receptor. Mol Endocrinol 28: 534–545. doi: 10.1210/me.2013-1359
|
| [18] | Mackerell AD, Feig M, Brooks CL (2004) Extending the treatment of backbone energetics in protein force fields: Limitations of gas-phase quantum mechanics in reproducing protein conformational distributions in molecular dynamics simulations. J Comp Chem 25: 1400–1415. |
| [19] |
Mackerell JAD, Bashford D, Bellott MLDR, et al. (1998) All-atom empirical potential for molecular modeling and dynamics studies of proteins. J Phys Chem B 102: 3586–3616. doi: 10.1021/jp973084f
|
| [20] |
Zoete V, Cuendet MA, Grosdidier A, et al. (2011) SwissParam: A fast force field generation tool for small organic molecules. J Comp Chem 32: 2359–2368. doi: 10.1002/jcc.21816
|
| [21] |
Humphrey W, Dalke A, Schulten K (1996) VMD: Visual molecular dynamics. J Mol Graph 14: 33–38. doi: 10.1016/0263-7855(96)00018-5
|
| [22] | Phillips JC, Braun R, Wang W, et al. (2005) Scalable molecular dynamics with NAMD. J Comp Chem 26: 1781–1802. |
| [23] |
Eftink MR, Anusiem AC, Biltonen RL (1983) Enthalpy-entropy compensation and heat capacity changes for protein-ligand interactions: General thermodynamic models and data for the binding of nucleotides to ribonuclease A. Biochemistry 22: 3884–3896. doi: 10.1021/bi00285a025
|
| [24] |
Sturtevant JM (1977) Heat capacity and entropy changes in processes involving proteins. Proc Natl Acad Sci 74: 2236–2240. doi: 10.1073/pnas.74.6.2236
|
| [25] |
Privalov PL, Potekhin SA (1986) Scanning microcalorimetry in studying temperature-induced changes in proteins. Method enzymol 131: 4–51. doi: 10.1016/0076-6879(86)31033-4
|
| [26] | Privalov PL, Dragan AI (2007) Microcalorimetry of biological macromolecules. Biophys Chem 126: 16–24. |
| [27] |
Gomez J, Hilser VJ, Xie D, et al. (1995) The heat capacity of proteins. Proteins: Struct, Funct, Bioinf 22: 404–412. doi: 10.1002/prot.340220410
|
| [28] |
Cooper A (2000) Heat capacity of hydrogen-bonded networks: An alternative view of protein folding thermodynamics. Biophys Chem 85: 25–39. doi: 10.1016/S0301-4622(00)00136-8
|
| [29] |
Pandey HD, Leitner DM (2017) Thermodynamics of hydration water around an antifreeze protein: A molecular simulation study. J Phys Chem B 121: 9498–9507. doi: 10.1021/acs.jpcb.7b05892
|
| [30] | Xu Y, Leitner DM (2014) Vibrational energy flow through the green fluorescent protein-water interface: Communication maps and thermal boundary conductance. J Phys Chem B 118: 7818–7826. |
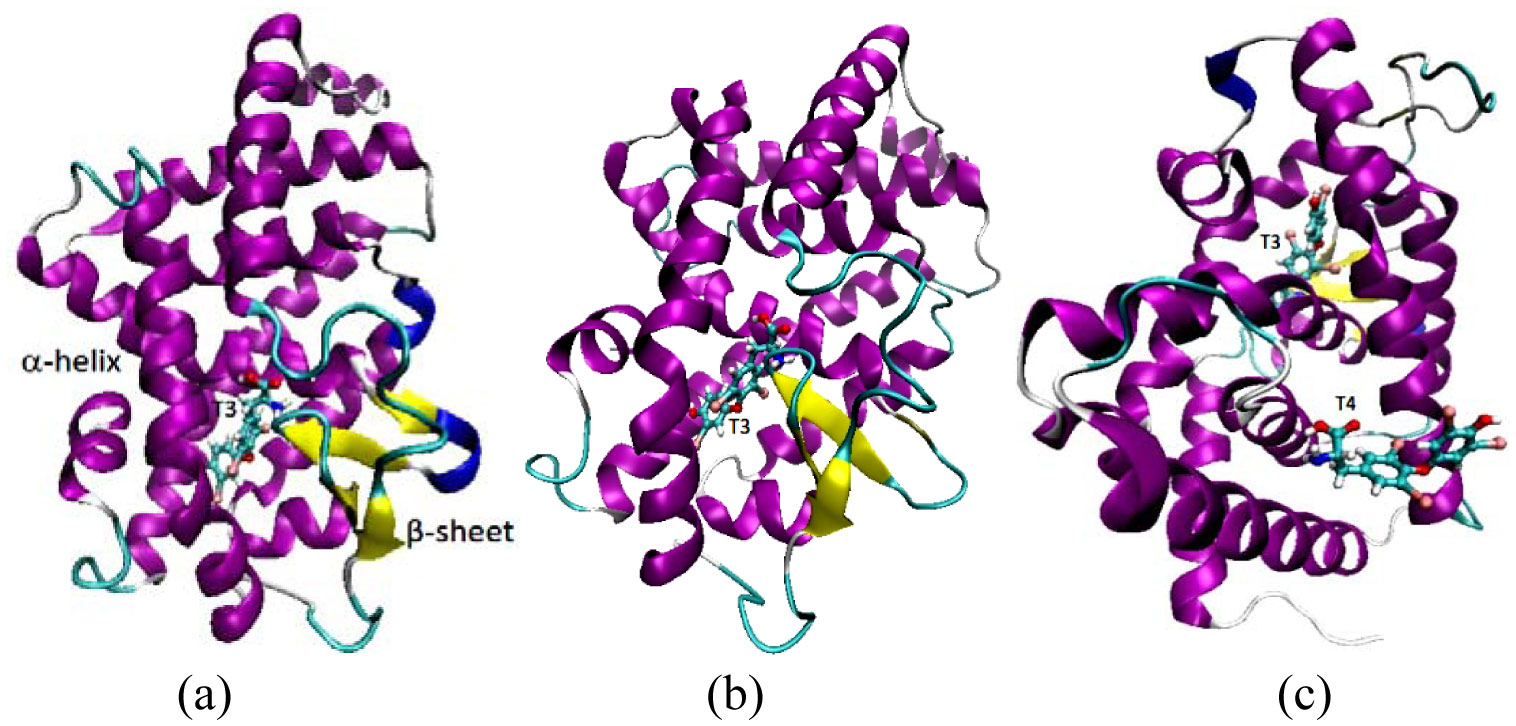
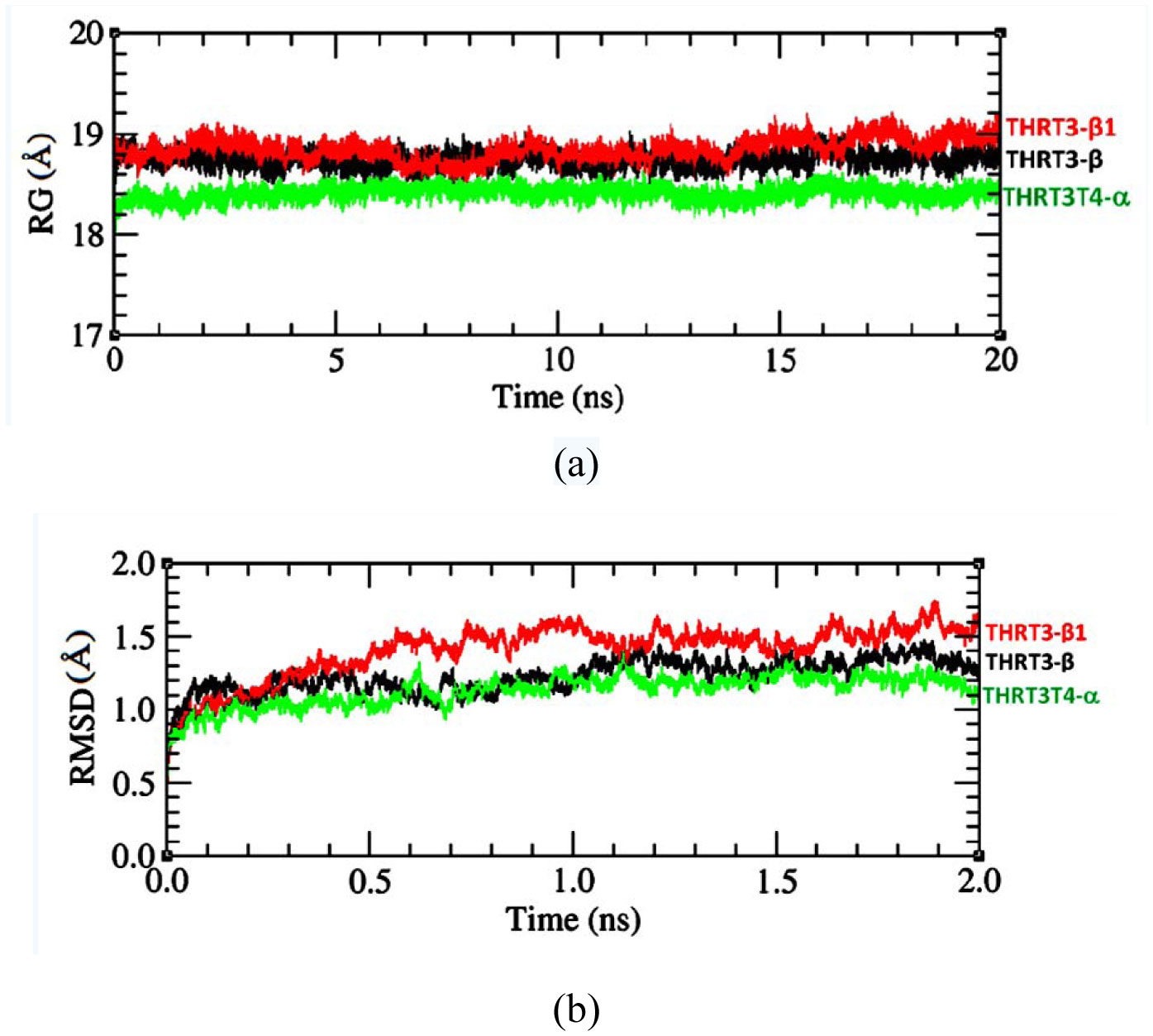
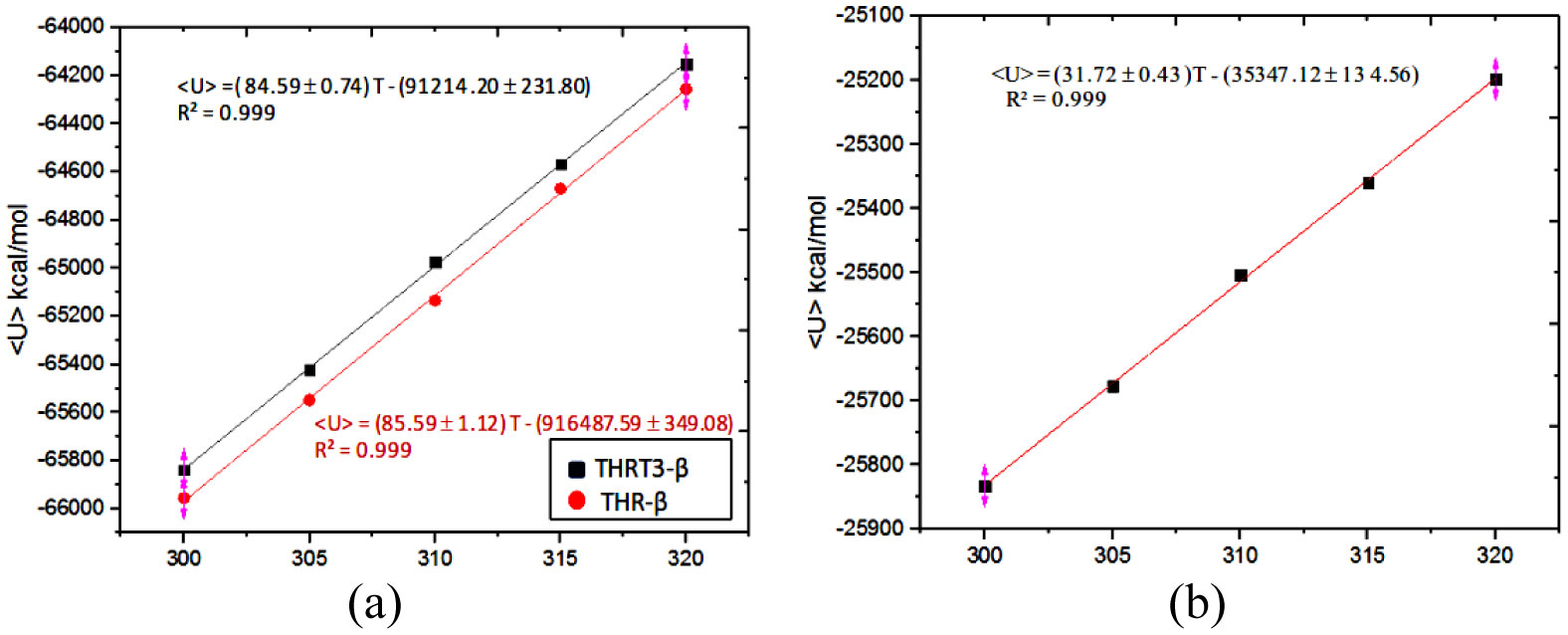
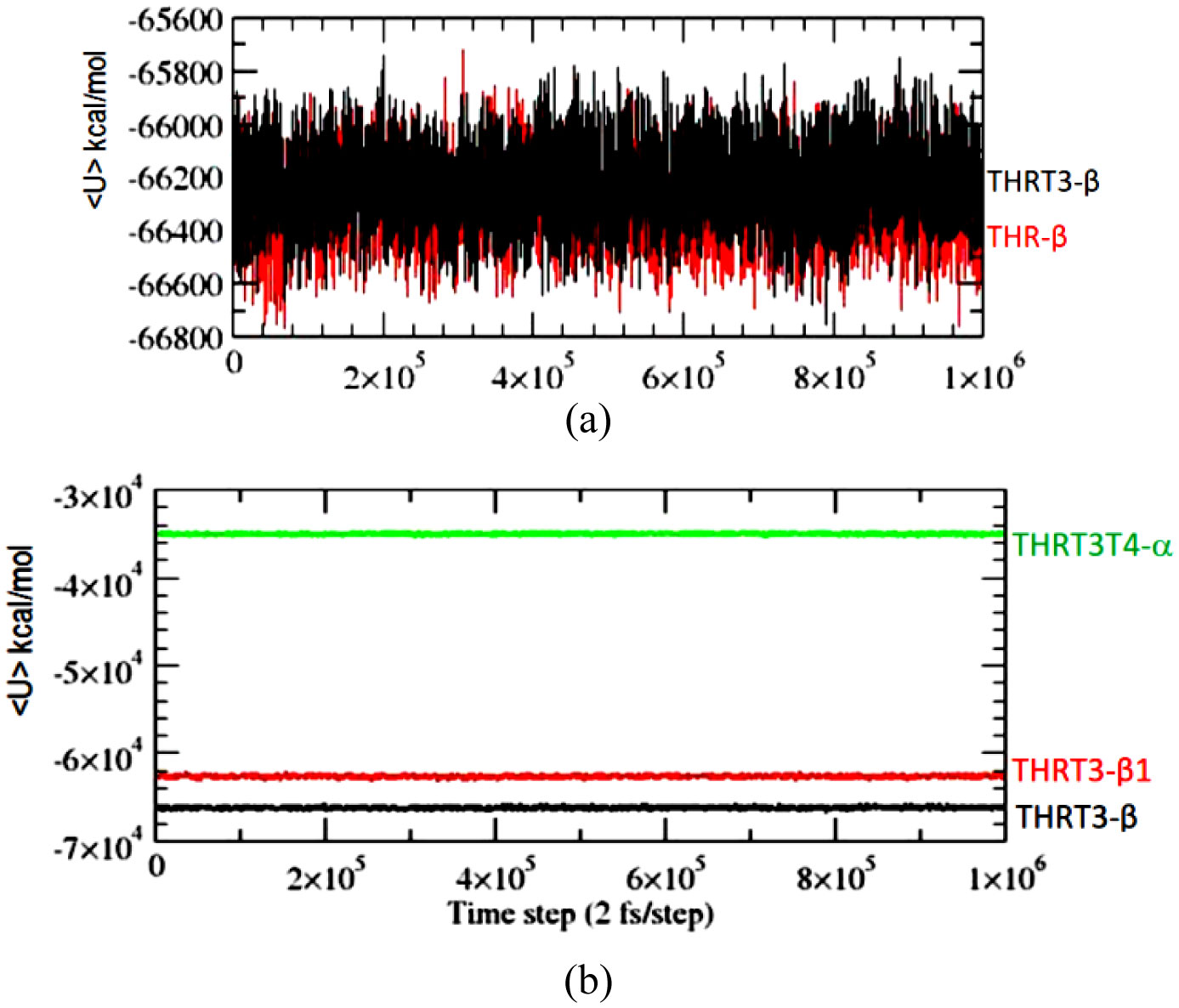
Figures(5) / Tables(2)

Tika Ram Lamichhane, Hari Prasad Lamichhane. Heat conduction by thyroid hormone receptors[J]. AIMS Biophysics, 2018, 5(4): 245-256. doi: 10.3934/biophy.2018.4.245







 DownLoad:
DownLoad: