




Figure 1.
The Graph G.
Citation: Elizabeth Ramírez-Iglesias, Rosa M. Hernández-Hernández, Carlos Bravo, José R. Ramírez-Iglesias, Pablo Herrera. Consumption, digestibility and cattle condition according to forage composition and quality in maize-cattle from conservation-based agroecosystems[J]. AIMS Agriculture and Food, 2020, 5(3): 480-499. doi: 10.3934/agrfood.2020.3.480
| [1] | Faten H. Damag, Amin Saif, Adem Kiliçman, Fozaiyah Alhubairah, Khaled M. Saad, Ekram E. Ali, Mouataz Billah Mesmouli . Monophonic sets and rough directed topological spaces: Applications with some directed networks. AIMS Mathematics, 2025, 10(8): 17623-17641. doi: 10.3934/math.2025787 |
| [2] | Hakeem A. Othman, Mohammed M. Al-Shamiri, Amin Saif, Santanu Acharjee, Tarik Lamoudan, Rashad Ismail . Pathless directed topology in connection to the circulation of blood in the heart of human body. AIMS Mathematics, 2022, 7(10): 18158-18172. doi: 10.3934/math.2022999 |
| [3] | R. Abu-Gdairi, A. A. El-Atik, M. K. El-Bably . Topological visualization and graph analysis of rough sets via neighborhoods: A medical application using human heart data. AIMS Mathematics, 2023, 8(11): 26945-26967. doi: 10.3934/math.20231379 |
| [4] | Ufuk Sevim, Leyla Goren-Sumer . Consensus of double integrator multiagent systems under nonuniform sampling and changing topology. AIMS Mathematics, 2023, 8(7): 16175-16190. doi: 10.3934/math.2023827 |
| [5] | Fawaz E. Alsaadi, Faisal Ali, Imran Khalid, Masood Ur Rehman, Muhammad Salman, Madini Obad Alassafi, Jinde Cao . Quantifying some distance topological properties of the non-zero component graph. AIMS Mathematics, 2021, 6(4): 3512-3524. doi: 10.3934/math.2021209 |
| [6] | Zeeshan Saleem Mufti, Ali Tabraiz, Qin Xin, Bander Almutairi, Rukhshanda Anjum . Fuzzy topological analysis of pizza graph. AIMS Mathematics, 2023, 8(6): 12841-12856. doi: 10.3934/math.2023647 |
| [7] | Ali N. A. Koam, Ali Ahmad, Azeem Haider, Moin A. Ansari . Computation of eccentric topological indices of zero-divisor graphs based on their edges. AIMS Mathematics, 2022, 7(7): 11509-11518. doi: 10.3934/math.2022641 |
| [8] | Usman Babar, Haidar Ali, Shahid Hussain Arshad, Umber Sheikh . Multiplicative topological properties of graphs derived from honeycomb structure. AIMS Mathematics, 2020, 5(2): 1562-1587. doi: 10.3934/math.2020107 |
| [9] | Shama Liaqat, Zeeshan Saleem Mufti, Yilun Shang . Newly defined fuzzy Misbalance Prodeg Index with application in multi-criteria decision-making. AIMS Mathematics, 2024, 9(8): 20193-20220. doi: 10.3934/math.2024984 |
| [10] | Gültekin Soylu, Müge Çerçi . Metrization of soft metric spaces and its application to fixed point theory. AIMS Mathematics, 2024, 9(3): 6904-6915. doi: 10.3934/math.2024336 |
Graph theory was introduced by Leonhard Euler for obtaining solution of the mathematical problem named "Seven Bridge of Königsberg" in 1736 [4]. The practices of the theory are used in the solution of many complex problems of modern life. Topology is an important branch of mathematics because of its contribution to the other branches of mathematics. Recently, the topology has been used as the appropriate frame for all sets connected by relations. Because rough sets and graphs are also based relational combinations, the topological structures of rough sets [2] and relation between rough sets and graphs are studied by some researchers [5,6,8].
An interesting research topic in graph theory is to study graph theory by means of topology. Some researches have created topologies from graphs using various methods. In 2013, M. Amiri et. al. have created a topology using vertices of an undirected graph [3]. In 2018, K.A. Abdu and A. Kılıçman have investigated the topologies generated by directed graphs [1].
In this paper, we aim at studying to create a topological space by using a simple undirected graph without isolated vertices. We present some properties of the topology that we create by using such graphs. We show that a topology can be generated by every simple undirected graph without isolated vertices. Moreover, we examine the topologies generated by using certain graphs. We define an equivalence relation on the set of the graphs with same vertices set. Finally, we give necessary and sufficient condition for continuity and openness of functions defined from one graph to another by using the topologies generated by these graphs. As a result of this, we present condition for the topological spaces generated by two different graphs to be homeomorphic.
In this section, some fundamental definitions and theorems related to the graph theory, approximation spaces and topological spaces used in the work are presented.
Definition 1. [4] A graph is an ordered pair of (U(G),E(G)), where U(G) is set of vertices, E(G) is set of edges linking to any unordered pair of vertices of G. If e is an edge linking to the vertices u and v, then it is said e links to vertices u and v. u and v called as ends of e. Moreover, it is said that these vertices are adjacent. A set of pairwise non-adjacent vertices of a graph is called an independent set. If the set of edges and vertices of a graph are finite, this graph is a finite graph. An edge whose ends are only one vertice is called a loop. An edge with distinct ends is a link.
Definition 2. [4] A graph is called simple graph, if there is at most one edge linking to arbitrary two vertices of the graph and it has not a loop.
Definition 3. [4] Let G=(U,E) be a graph. If vertices set U can divided into two subsets A and B so that each edge of G has one end in A and one end in B, G is called bipartite graph. In other words, a graph G is bipartite iff vertices set U of G can divided into two independent sets.
Definition 4. [4] A walk is a sequence of finite number of adjacent vertices such that v0e1v1e2v2...ekvk in a graph G=(U,E). A walk that each edge and vertice is used at most one time is called a path.
Definition 5. [4] A cycle is a path with the same starting and ending point and it is denoted with Cn.
Theorem 1. [7] Let X be a nonempty set and β be a class of subsets of X. If following conditions are satisfied, the collection β is a base just for one topology.
1. X=⋃B∈βB
2. For B1∈β and B2∈β, the set B1∩B2 is union of some set belonging to β.
Definition 6. Let G=(U,E) be a graph. Then the set of vertices becoming adjacent to a vertice u is called adjacency of u and it is denoted AG(u). Minimal adjacency of u is defined as
| [u]G=⋂u∈AG(v)AG(v). |
Theorem 2. Let G=(U,E) be a simple undirected graph without isolated vertices. Then the class βG={[u]G:u∈U} is a base for a topology on U.
Proof. Firstly, we shall show that ⋃u∈U[u]G=U. From definition of [u]G, u∈[u]G is obtained for every u∈U. Since the graph G is a graph without isolated vertices, the class {[u]G:u∈U} covers to the set U. That is,
| ⋃u∈U[u]G=U. |
Secondly, we shall show that there exists V⊆U such that [u]G∩[v]G=⋃w∈V⊆U[w]G, for every [u]G, [v]G∈βG. Let [u]G,[v]G∈βG. Then [u]G∩[v]G=∅ or [u]G∩[v]G≠∅. If [u]G∩[v]G=∅, it is seen that [u]G∩[v]G=⋃w∈∅[w]G since ⋃w∈∅[w]G=∅. If [u]G∩[v]G≠∅, there exists at least one w∈U such that w∈[u]G∩[v]G. Then w belongs to both [u]G and [v]G. Since w∈[u]G, it is seen that w∈AG(t), for all t∈U such that u∈AG(t). Similarly, since w∈[v]G, it is seen that w∈AG(t′), for all t′∈U such that v∈AG(t′).
Hence, w∈AG(t) and w∈AG(t′)⇒w∈AG(t)∩AG(t′)
| ⇒⋃w∈AG(t)∩AG(t′)[w]G (Since w∈[w]G)⇒w∈⋃w∈V⊆U[w]G. (V=AG(t)∩AG(t′)) |
Then it is obtained that
| [u]G∩[v]G⊆⋃w∈V⊆U[w]G | (3.1) |
On the other hand,
| k∈⋃w∈AG(t)∩AG(t′)[w]G⇒k∈[w]G, for ∃w∈AG(t)∩AG(t′) |
| ⇒k∈⋂w∈AG(w′)AG(w′)⇒k∈AG(w′), for all w∈AG(w′)⇒k∈AG(t)∩AG(t′)⇒k∈⋂u∈AG(t)AG(t) and k∈⋂v∈AG(t′)AG(t′)⇒k∈[u]G and k∈[v]G⇒k∈[u]G∩[v]G. |
Then it is obtained that
| ⋃w∈AG(t)∩AG(t′)[w]G⊆[u]G∩[v]G. | (3.2) |
Therefore, the following equation is obtained from (3.1) and (3.2):
| [u]G∩[v]G=⋃w∈AG(t)∩AG(t′)[w]G. |
Consequently, βG is a base for a topology on U.
Corollary 1. Each simple undirected graph without isolated vertices creates a topology on vertices set of the graph.
Definition 7. Let G=(U,E) be a simple undirected graph without isolated vertices. Then the topology generated by βG={[u]G:u∈U} is called the topology generated by the graph G. This topology is in the form of:
| τG={G⊆U:G=⋃[u]G∈βG[u]G,u∈V⊆U}. |
Here, the class of closed sets of this topology is in the form of:
| KG={Gc:G∈τG}. |
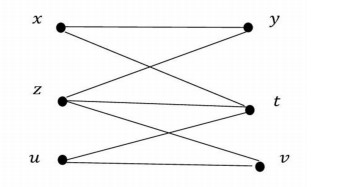
Example 1. The graph whose vertices set is U={x,y,z,t,u,v} is given in Figure 1.
The minimal adjacencies of each vertice are as follows:
| [x]G={x,z},[y]G={y,t},[z]G={z},[t]G={t},[u]G={z,u},[v]G={t,v}. |
Thus,
| βG={{z},{t},{x,z},{y,t},{z,u},{t,v}} |
and
| τG={U,∅,{z},{t},{x,z},{y,t},{z,u},{t,v},{z,t},{y,z,t},{z,t,v},{x,z,t},{z,t,u},{x,y,z,t},{x,z,u},{x,z,t,v},{y,z,t,u},{y,t,v},{x,z,t,u},{x,y,z,t,u},{x,y,z,t,v},{y,z,t,v},{z,t,u,v},{x,z,t,u,v},y,z,t,u,v}}. |
τG is topology generated by G. The class of closed sets of this topology is
| KG={U,∅,{x,y,t,u,v},{x,y,z,u,v},{y,t,u,v},{x,z,u,v},{x,y,t,v},{x,y,z,u},{x,y,u,v},{x,u,v},{x,y,u},{y,u,v},{x,y,v},{u,v},{y,t,v},{y,u},{x,v},{x,z,u},{y,v},{v},{u},{x,u},{x,y},{y},{x}}. |
Class of both open and closed sets is as follows:
| CO(U)={U,∅,{x,z,u},{y,t,v}}. |
Here, it is seen that both open and closed sets different from U and ∅ are {x,z,u} and {y,t,v}. Moreover, the graph G is bipartite and these sets are independent sets whose intersection is ∅ and union is U.
Theorem 3. Let KA,B=(U,E) be a complete bipartite graph. Then the topology generated by KA,B is a quasi-discrete topology.
Proof. Since KA,B is a bipartite graph, A∩B=∅ and A∪B=U. For every x∈U, x∈A or x∈B. Let x∈A. Since KA,B is a complete bipartite graph, we have AKA,B(x)=B and [x]KA,B=A. Let x∈B. Then we have AKA,B(x)=A and [x]KA,B=B. Hence, the base of the topology generated by KA,B is as follows:
| βKA,B={A,B}. |
Therefore, the topology generated by KA,B is as follows:
| τKA,B={A,B,∅,U}. |
τKA,B is a quasi-discrete topology on U.
Theorem 4. Let Kn=(U,E) be a complete graph, where U={v1,v2,...,vn}. Then the topology generated by Kn is discrete topology on U.
Proof. The minimal neighborhoods of vertices set U={v1,v2,...,vn} are as follows respectively:
| [v1]G={v1},[v2]G={v2},...,[vn]G={vn}. |
Therefore,
| βKn={{vn}:vn∈U} |
and the topology generated by Kn is as follows:
| τKn=P(U). |
It is seen that τKn is discrete topology on U.
Example 2. Let us investigate the topological space generated by C5 given Figure 2 whose vertices set is U={v1,v2,v3,v4,v5}.
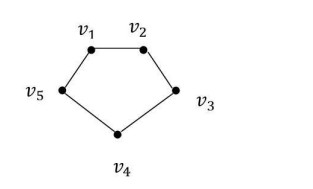
The adjacencies of the vertices of the cycle C5 are as follows:
| AG(v1)={v2,v5},AG(v2)={v1,v3},AG(v3)={v2,v4},AG(v4)={v3,v5},AG(v5)={v1,v4}. |
The minimal adjacencies of the vertices of the cycle C5 are as follows:
| [v1]G=⋂v1∈AG(u)AG(u)={v1},[v2]G={v2},[v3]G={v3},[v4]G={v4},[v5]G={v5}. |
Thus,
| βC5={{v1},{v2},{v3},{v4},{v5}}. |
The class βC5 is a base for the discrete topology on U. Thus, the topological space generated by this graph is discrete topological space on U.
Theorem 5. Let Cn=(U,E) be a cycle whose vertices set is U={v1,v2,...,vn}, where n≥3 (n≠4). Then the topological space generated by the cycle Cn=(U,E) is a discrete topological space.
Proof. The graph Cn is as in Figure 3.
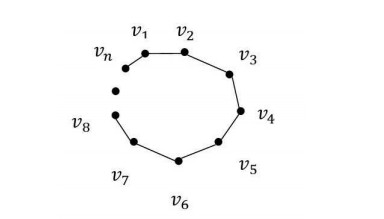
The adjacencies of the vertices of the cycle Cn are as follows:
| AG(v1)={vn,v2},AG(v2)={v1,v3},AG(v3)={v2,v4},...,AG(vn−1)={vn−2,vn},AG(vn)={vn−1,v1}. |
The minimal adjacencies of the vertices of the cycle Cn are as follows:
| [v1]G=⋂v1∈AG(vi)AG(vi)={v1},[v2]G=⋂v2∈AG(vi)AG(vi)={v2},...,[vn−1]G=⋂vn−1∈AG(vi)AG(vi)={vn−1},[vn]G=⋂vn∈AG(vi)AG(vi)={vn}. |
Thus, we have
| βCn={{vn}:Vn∈U}. |
The class βCn is a base for discrete topology on U. Thus, it is seen that the topological space generated by the graph Cn is the discrete topological space on U.
When we assume n=4, the graph C4 is a complete bipartite graph. The topological space generated the graph C4 whose vertices set is U={v1,v2,v3,v4} is τC4={U,∅,{v1,v4},{v2,v3}}. This topology is not discrete topology, but it is quasi-discrete topology.
Remark 1. Two different graph G and G′with same vertices set can create the same topology. It is seen clearly that although the graphs Kn and Cn with same vertices set is different these graphs create same topology.
Theorem 6. Let G be set of all simple undirected graphs whose vertices set is U={v1,v2,...,vn} without isolated vertices. The relation ∼ defined on G as "G1∼G2⇔τG1=τG2" is a equivalence relation.
Proof. ⅰ) Since τG1=τG1,G1∼G1
ⅱ) Let G1∼G2.From definition "∼", it is seen that τG1=τG2. Since τG2=τG1, we obtain that G2∼G1.
ⅲ) Let G1∼G2 and G2∼G3. Then it is seen that τG1=τG2and τG2=τG3. Thus, we obtain that τG1=τG3. Consequently, we obtain that G1∼G3.
Since G is symmetric, transitive and reflexive, it is an equivalence relation.
Theorem 7. Let G=(U,E) and G′=(U′,E′)\ be two graphs without isolated vertices. Let τG and τG′ be the topologies generated by G and G′ respectively and f:(U,τG)→(U′,τG′) be a function. Then f is continuous iff for every u∈U,
| f([u]G)⊆[f(u)]G′. |
Proof. Let f:(U,τG)→(U′,τG′) be a continuous function. Then βG={[u]G:u∈U} and βG′={[u′]G′:u′∈U′} are bases of topologies τG and τG′, respectively. Since f is continuous, there is B∈βG such that f(B)⊆[f(u)]G′ for every u∈U. [u]G is the minimal element containing u of βG. Thus, it is obtained that
| f([u]G)⊆[f(u)]G′. |
Conversely, let f([u]G)⊆[f(u)]G′ for every u∈U. It is seen that [f(u)]G′∈βG′f(v), for every u∈U. Since f([u]G)⊆[f(u)]G′ and [u]G∈βGv, the function f is a continuous function.
Example 3. Let us investigate the graphs G=(U,E) and G′=(U′,E′) is given in Figure 4. Let f:(U,τG)→(U′,τG′) be a function defined by
| f(x)=f(z)=a,f(l)=b,f(y)=c,f(t)=d,f(k)=e. |
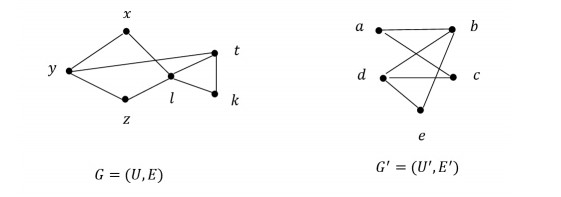
The minimal adjacencies of vertices of G are follows:
| [x]G={x,z},[y]G={y,l},[z]G={x,z},[t]G={t},[l]G={l},[k]G={k}. |
The minimal adjacencies of vertices of G′ are as follows:
| [a]G′={a,d},[b]G′={b},[c]G′={b,c},[d]G′={d},[e]G′={e}. |
It is seen that f([v]G)⊆[f(v)]G′, for every v∈U. Therefore, f is a continuous function.
Corollary 2. Let f:(U,τKn)→(U′,τG′) be arbitrary function, where Kn=(U,E) is a complete graph and G′=(U′,E′) is arbitrary graph. Then f is continuous function.
Theorem 8. Let G=(U,E) and G′=(U′,E′) be two simple undirected graphs without isolated vertice and τG and τG′ the topologies generated by this graphs, respectively. Let f:(U,τG)→(U′,τG′) be a function. Then f is open function iff for every u∈U,
| [f(u)]G′⊆f([u]G). |
Proof. Let f:(U,τG)→(U′,τG′) be an open function. Then f([u]G) is an open subset of U′ for every [u]G∈βG. It is obtained that
| f(u)∈[f(u)]G′⊆f([u]G). |
Therefore, we have
| [f(u)]G′⊆f([u]G). |
Conversely, Let [f(u)]G′⊆f([u]G), for every u∈U. It is seen that for every u∈U,
| f(u)∈[f(u)]G′⊆f([u]G). |
Thus, f([u]G) is an open subset of U′. Consequently, we can say f is an open function.
From above theorem, it is seen that an open function may not continuous and a continuous function may also not be open. Now we give a necessary and sufficient condition for a function to be continuous and open.
Theorem 9. Let G=(U,E) and G′=(U′,E′) be simple undirected two graphs without isolated vertices and τG and τG′ the topologies generated by this graphs, respectively. Let f:(U,τG)→(U′,τG′) be a function. Then f is a open and continuous function iff for every u∈U,
| [f(u)]G′=f([u]G). |
Proof. It is clearly seen from Theorem 3.6 and Theorem 3.7.
Corollary 3. Let G=(U,E) and G′=(U′,E′) be simple undirected two graphs without isolated vertice and τG and τG′the topologies generated by this graphs, respectively. Let f:(U,τG)→(U′,τG′)\ be a function. Then f is a homeomorphism iff f is a bijection that for every u∈U,
| [f(u)]G′=f([u]G). |
In this paper it is shown that topologies can be generated by simple undirected graphs without isolated vertices. It is studied topologies generated by certain graphs. Therefore, it is seen that there is a topology generated by every simple undirected graph without isolated vertices. Properties proved by these generated topologies are presented. An equivalence of the graphs with same vertices set is defined. Finally, necessary and sufficient condition is given for continuity and openness of a function defined to another graph from one graph. This enables us to determine whether these topological spaces is homeomorphic without needing to find the topological spaces generated by two graphs.
The first author would like to thank TUBITAK (The Scientific and Technological Research Council of Turkey) for their financial supports during her PhD studies.
The authors declare that they have no competing interest.
| [1] | Herrera P, Birbe B, Colmenares O, et al. (2008) Sistemas de producción con ganadería de doble propósito en condiciones de sabanas bien drenadas. Acta Biol Venez 28: 29-38. |
| [2] | Mendoza JC, Aparicio YV, Jerez MP, et al. (2013) Ecological evaluation of three agroecosystems of sheep production in Central Valleys of Oaxaca. Rev Mex Cienc Agr 6: 1251-1261. |
| [3] | Clements RJ (1996) Pastures for prosperity. 3. The future for new tropical pasture plants. Trop Grasslands 30: 31-46. |
| [4] |
Nakamura T, Miranda CH, Ohwaki Y, et al. (2005) Characterization of nitrogen utilization by Brachiaria grasses in Brazilian Savannas (Cerrados). Soil Sci Plant Nutr 51: 973-979. doi: 10.1111/j.1747-0765.2005.tb00136.x

|
| [5] | Vercoe JE (1996) Pastures for prosperity. 5. The role of animal science. Trop Grasslands 30: 58-72. |
| [6] | Hernández-Hernández RM, Lozano Z, Toro M, et al. (2011) Informe final de proyecto: "Manejo agroecológico de suelos de sabanas bien drenadas con unidades de producción cereal-ganado." FONACIT-MPPCyT. Caracas, Venezuela, 202. |
| [7] |
Benvenutti MA, Pavetti DR, Poppi DP, et al. (2016) Defoliation patterns and their implications for the management of vegetative tropical pastures to control intake and diet quality by cattle. Grass Forage Sci 71: 424-436. doi: 10.1111/gfs.12186
|
| [8] | Barriga J, Visbal M, Acero J (2011) Relación entre los caracteres de las micorrizas arbusculares nativas con las propiedades físico-químicas del suelo y bromatología del pasto estrella en ganadería de carne. Rev Científica UDO Agr 11: 134-141. |
| [9] |
Pedreira CG, Braga GJ, Portela JN (2017) Herbage accumulation, plant-part composition and nutritive value on grazed signal grass (Brachiaria decumbens) pastures in response to stubble height and rest period based on canopy light interception. Crop Pasture Sci 68: 62-73. doi: 10.1071/CP16333
|
| [10] |
Trevisi E, Riva F, Filipe JFS, et al. (2018) Innate immune responses to metabolic stress can be detected in rumen fluids. Res Vet Sci 117: 65-73. doi: 10.1016/j.rvsc.2017.11.008
|
| [11] |
Cuchillo-Hilario M, Wrage-Mönnig N, Isselstein J (2018) Forage selectivity by cattle and sheep co-grazing swards differing in plant species diversity. Grass Forage Sci 73: 320-329. doi: 10.1111/gfs.12339
|
| [12] |
Couto RK, Pinto GG, Rodrigues R, et al. (2016) Comparison of protein and energy supplementation to mineral supplementation on feeding behavior of grazing cattle during the rainy to the dry season transition. Springerplus 5: 2-7. doi: 10.1186/s40064-015-1651-x
|
| [13] |
Maestri R, Monteiro LR, Fornel R, et al. (2018) Geometric morphometrics meets metacommunity ecology: environment and lineage distribution affects spatial variation in shape. Ecography 41: 90-100. doi: 10.1111/ecog.03001
|
| [14] | Anza M, Epelde L, Mijangos I, et al. (2016) Effects of grazing abandonment on the health of pasture ecosystems. Pastos 45: 15-22. |
| [15] |
Vélez OM, Campos R, Sánchez H, et al. (2017) Evaluación de diferentes niveles de inclusión de plantas nativas de sabanas inundables sobre una dieta basal de Brachiaria humidicola y su efecto sobre la producción de metano in vitro. Arch Zootec 66: 341-349. doi: 10.21071/az.v66i255.2509
|
| [16] |
DePeters EJ, George LW (2014) Rumen transfaunation. Immunol Lett 162: 69-76. doi: 10.1016/j.imlet.2014.05.009
|
| [17] | Smith LW, Goering HK, Gordon CH (1969) Influence of chemical treatments upon digestibility of ruminant feces. Proc Conf on Anim Waste Manage 88-97. |
| [18] | Caballero JC, Moreno A, Reyes JL, et al. (2017) Competencia del uso del rastrojo de maíz en sistemas agropecuarios mixtos en Chiapas. Rev Mex Cienc Agr 8: 91-104. |
| [19] |
Orgill SE, Condon JR, Conyers MK, et al. (2018) Removing grazing pressure from a native pasture decreases soil organic carbon in southern New South Wales, Australia. Land Degrad Dev 29: 274-283. doi: 10.1002/ldr.2560
|
| [20] | Ormaechea S, Gargaglione V, Bahamonde HA, et al. (2018) Producción bovina bajo manejo silvopastoril intensivo a escala de establecimiento y ciclo completo en Tierra del Fuego, Argentina. Livest Res Rural Dev 30: 1-12. |
| [21] | Gonçalves Z, Junqueira EC, Pinto F, et al. (2019) Morphogenic, structural, productive and bromatological characteristics of Braquiária in silvopastoral system under nitrogen doses. Acta Sci Anim Sci 41: 1-8. |
| [22] | Santos BRC dos, Alfaya H, Dias AEA, et al. (2011) Correlation of the phosphorus in the system soil-plant-animal in natural pasture in the region of campanha-rs. Arch Zootec 60: 487-497. |
| [23] | Givens DI, Owen E, Omed HM, et al. (2000) Forage Evaluation in Ruminant Nutrition. CABI, Wallingford, UK, 498. |
| [24] | Ginantra IK, Made IB, Muksin IK (2018) Selection of forages by timor deer (Cervus timorensis blainville) in Menjangan island, Bali. IOP Conf Ser Earth Environ Sci 130: 1-12. |
| [25] | Sparks DR, Malechek JC (1968) Estimating percentage dry weight in diets using a microscopic technique. Rangeland Ecol Manag/J Range Manag Arch 21: 264-265. |
| [26] |
Tóth E, Deák B, Valkó O, et al. (2018) Livestock type is more crucial than grazing intensity: traditional cattle and sheep grazing in short-grass steppes. Land Degrad Dev 29: 231-239. doi: 10.1002/ldr.2514
|
| [27] | Mata D, Herrera P, Birbe B (1996) Sistemas de producción animal con bajos insumos para las sabanas de Trachypogon sp. Ecotropicos 9: 83-100. |
| [28] | Ramia M (1967) Tipos de sabanas en los llanos de Venezuela. Bol Soc Ven Cienc Nat 27: 264-288. |
| [29] | Hernández-Hernández RM, Florentino A, López-Hernández D (2000) Efectos de la siembra directa y la labranza convencional sobre la estabilidad estructural y otras propiedades físicas de un suelo de sabana. Agron Trop 50: 9-29. |
| [30] | Carpenter SR (1998) The Need for Large-Scale Experiments to Assess and Predict the Response of Ecosystems to Perturbation, In: Pace ML, Groffman PM, editors. Successes, Limitations, and Frontiers in Ecosystem Science, New York, NY: Springer, 287-312. |
| [31] | Lozano Z, Bravo C, Ovalles F, et al. (2004) Selección de un diseño de muestreo en parcelas experimentales a partir del estudio de la variabilidad espacial de los suelos. Bioagro 16: 61-72. |
| [32] | AOAC (1998) Methods of analysis (16th). Washington D.C. EUA. |
| [33] | Brown S (1997) Estimating Biomass and Biomass Change of Tropical Forests: A Primer. FAO Forestry Paper, Roma, Italy, 134. |
| [34] | Sepúlveda L, Pelliza A, Manacorda M (2004) La importancia de los tejidos no epidérmicos en el microanálisis de la dieta de herbívoros. Ecol Austral 14: 31-38. |
| [35] | Castellaro G, Ullrich T, Wackwitz B, et al. (2004) Composición botánica de la dieta de alpacas (Lama pacos L.) y llamas (Lama glama L.) en dos estaciones del año, en praderas altiplánicas de un sector de la Provincia de Parinacota, Chile. Agr Técnica 64: 353-363. |
| [36] |
Holechek JL, Vavra M, Pieper RD (1982) Botanical composition determination of range herbivore diets: a review. J Range Manag 35: 309-315. doi: 10.2307/3898308
|
| [37] |
Morante N, Sánchez T, Ceballos H, et al. (2010) Tolerance to postharvest physiological deterioration in cassava roots. Crop Sci 50: 1333-1338. doi: 10.2135/cropsci2009.11.0666
|
| [38] | Soest PJV (2018) Nutritional Ecology of the Ruminant. Ithaca, NY: Cornell University Press, 490. |
| [39] | Pardo O, Carulla JE, Hess HD (2008) Efecto de la relación proteína y energía sobre los niveles de amonio ruminal y nitrógeno ureico en sangre y leche de vacas doble propósito del piedemonte llanero, Colombia. Rev Col Cienc Pecuarias 21: 387-397. |
| [40] | Di Rienzo JA, Casanoves F, Balzarini MG, et al. (2011) Grupo InfoStat FCA, Universidad Nacional de Córdoba, Argentina. 8: 195-199. |
| [41] | Uriel E, Manzano JA (2002) Análisis multivariante aplicado. Paraninfo 76: 270-271. |
| [42] | Ramírez-Iglesias E, Hernández-Hernández RM, Herrera P (2017) Relaciones suelo-planta-animal en un sistema agroecológico de siembra directa y asociación de coberturas maíz-ganado en sabanas bien drenadas de Venezuela. Acta Biol Venez 37: 67-87. |
| [43] | Rodas-González A, Vergara-López J, Arenas L, et al. (2006) Características al sacrificio, rasgos de la canal y rendimiento carnicero de novillos criollo limonero sometidos a suplementación durante la fase de ceba a pastoreo. Rev Científica 16: 315-324. |
| [44] |
Carvalho ÁC, Pereira M, Bonomo P, et al. (2014) Microbial protein synthesis and nitrogen metabolism in cows bred on tropical pasture and fed on cassava root and corn. Acta Sci Anim Sci 36: 185-192. doi: 10.4025/actascianimsci.v36i2.22161
|
| [45] | Russell JB, O'Connor JD, Fox DG, et al. (1992) A net carbohydrate and protein system for evaluating cattle diets: I. Ruminal fermentation. J Anim Sci 70: 3551-3561. |
| [46] |
Pfeiffer M, Langan L, Linstädter A, et al. (2019) Grazing and aridity reduce perennial grass abundance in semi-arid rangelands - Insights from a trait-based dynamic vegetation model. Ecol Model 395: 11-22. doi: 10.1016/j.ecolmodel.2018.12.013
|
| [47] | Sevim Ö, Önol AG (2019) Supplemental slow-release urea and non-structural carbohydrates: effect on digestibility and some rumen parameters of sheep and goats. J Anim Plant Sci 29: 1-7. |
| [48] | Ramírez-Iglesias E, Hernández-Hernández RM, Herrera P (2017) Dinámica del fósforo en un agroecosistema maíz-ganado de sabanas neotropicales. Rev Col Cienc Anim Recia 9: 147-157. |
| [49] | La O, González H, Orozco A, et al. (2012) Composición química, degradabilidad ruminal in situ y digestibilidad in vitro de ecotipos de Tithonia diversifolia de interés para la alimentación de rumiantes. Rev Cubana Cienc Agr 46: 47-53. |
| [50] | Singh S, Nag SK, Kundu SS, et al. (2010) Relative intake, eating pattern, nutrient digestibility, nitrogen metabolism, fermentation pattern and growth performance of lambs fed organically and inorganically produced cowpea hay- barley grain diets. Trop Grasslands 44: 55-61. |
| [51] | Giráldez FJ, Mantecón ÁR, Chaso MA, et al. (1994) Efecto del tipo de dieta y de la frecuencia de alimentación sobre la actividad degradativa ruminal. Prod Sanid Anim 9: 245-259. |
| [52] |
Satter LD, Slyter LL (1974) Effect of ammonia concentration on rumen microbial protein production in vitro. British J Nutr 32: 199-208. doi: 10.1079/BJN19740073
|
| [53] | Chamorro DR, Carulla JE, Cuesta PA (2005) Caracterización nutricional de dos asociaciones gramínea-leguminosa con novillas en pastoreo en el Alto Magdalena. Ciencia y Tecnología Agropecuaria 6: 37-51. |
| [54] | Sales E, Varela AM, Ferreira L, et al. (2005) Importância da inter-relação carboidrato e proteína em dietas de ruminantes. Sem Ci Agr 30: 125-134. |
| [55] | Bravo C, Lozano Z, Hernández-Hernández RM, et al. (2004) Efecto de diferentes especies de coberturas sobre las propiedades físicas de un suelo de sabana con siembra directa de maíz. Bioagro 16: 163-172. |
| [56] | Lozano Z, Hernández-Hernández RM, Bravo C, et al. (2012) Disponibilidad de fósforo en un suelo de las sabanas bien drenadas venezolanas, bajo diferentes tipos de cobertura y fuentes de fertilización. Interciencias 37: 820-827. |
| [57] | Canelones C, Castejon M (2006) Flours of whole cowpea (Vigna unguiculata) and cob maize (Zea mays) as supplements for suckling calves. Zootec Trop 24: 361-378. |
| [58] | Milligan KE, Brookes IM, Thompson KF (1987) Feed planning on pasture, In: Nicol, A.M., Livestock feeding on pasture, New Zealand Society of Animal Production: Hamilton, 75-88. |
| [59] | Thiago LRL, Gill M (1990) Consumo voluntário: fatores relacionados com a degradação e passagem da forragem pelo rumen. EMBRAPA, Campo Grande, Brasil, 65. |
| 1. | Abdulgani Şahin, Gohar Ali, Further Results on Total Edge-Vertex Domination, 2022, 2022, 2314-4785, 1, 10.1155/2022/5439610 | |
| 2. | Hakeem A. Othman, Mohammed M. Al-Shamiri, Amin Saif, Santanu Acharjee, Tarik Lamoudan, Rashad Ismail, Pathless directed topology in connection to the circulation of blood in the heart of human body, 2022, 7, 2473-6988, 18158, 10.3934/math.2022999 | |
| 3. | B K. Mahmoud, Y Y Yousif, Compatibility and Edge Spaces in Alpha - Topological Spaces, 2021, 1963, 1742-6588, 012073, 10.1088/1742-6596/1963/1/012073 | |
| 4. | Nechervan B. Ibrahim, Alias B. Khalaf, On maximal block topological space, 2023, 45, 10641246, 8541, 10.3233/JIFS-223749 | |
| 5. | Hakeem Othman, Ahmed Ayache, Amin Saif, On L2−directed topological spaces in directed graphs theory, 2023, 37, 0354-5180, 10005, 10.2298/FIL2329005O | |
| 6. | Faten H. Damag, Amin Saif, Adem Kiliçman, Ekram E. Ali, Mouataz B. Mesmouli, On m-Negative Sets and Out Mondirected Topologies in the Human Nervous System, 2024, 12, 2227-7390, 3763, 10.3390/math12233763 | |
| 7. | Faten H. Damag, Amin Saif, Adem Kiliçman, Mouataz Billah Mesmouli, Fozaiyah Alhubairah, Upper a-Graphical Topological Spaces with the COVID-19 Form and Its Diffusion, 2025, 14, 2075-1680, 84, 10.3390/axioms14020084 | |
| 8. | Husniyah Alzubaidi, Ljubiša D. R. Kočinac, Hakeem A. Othman, On Topologies on Simple Graphs and Their Applications in Radar Chart Methods, 2025, 14, 2075-1680, 178, 10.3390/axioms14030178 | |
| 9. | Quang-Thinh Bui, Thanh Nha Nguyen, Tzung-Pei Hong, Bay Vo, Graph-induced topological space: from topologies to separation axioms, 2025, 1432-7643, 10.1007/s00500-025-10633-9 | |
| 10. | Faten H. Damag, Amin Saif, Adem Kiliçman, Fozaiyah Alhubairah, Khaled M. Saad, Ekram E. Ali, Mouataz Billah Mesmouli, Monophonic sets and rough directed topological spaces: Applications with some directed networks, 2025, 10, 2473-6988, 17623, 10.3934/math.2025787 |

Figures(5) / Tables(3)

Elizabeth Ramírez-Iglesias, Rosa M. Hernández-Hernández, Carlos Bravo, José R. Ramírez-Iglesias, Pablo Herrera. Consumption, digestibility and cattle condition according to forage composition and quality in maize-cattle from conservation-based agroecosystems[J]. AIMS Agriculture and Food, 2020, 5(3): 480-499. doi: 10.3934/agrfood.2020.3.480

 DownLoad:
DownLoad:
