Citation: Giuseppina Albano, Virginia Giorno. Inference on the effect of non homogeneous inputs in Ornstein-Uhlenbeck neuronal modeling[J]. Mathematical Biosciences and Engineering, 2020, 17(1): 328-348. doi: 10.3934/mbe.2020018

| [1] | G. L. Gerstein and B. Mandelbrot, Random walk models for the spike activity of a single neuron, Biophys. J., 4 (1964), 41-68. |
| [2] | H. C. Tuckwell, Introduction to Theoretical Neurobiology 2. Nonlinear and Stochastic Theories, Cambridge Univ. Press, Cambridge, 1988. |
| [3] | S. Ditlevsen and P. Lánský, Estimation of the input parameters in the Ornstein-Uhlenbeck neuronal ´ model, Phys. Rev. E, 71 (2005), 011907. |
| [4] | P. Lánský, P. Sanda and J. He, The parameters of the stochastic leaky integrate-and-fire neuronal model, J. Comput. Neurosci., 21 (2006), 211-223. |
| [5] | S. Ditlevsen and P. Lánský, Comparison of statistical methods for estimation of the input parameters in the Ornstein-Uhlenbeck neuronal model from first-passage times data, in: "American Institute of Physics Proceedings Series, CP1028, Collective Dynamics: Topics on Competition and Cooperation in the Biosciences", Eds.: L.M. Ricciardi, A. Buonocore, and E. Pirozzi, (2008), 171-185. |
| [6] | E. Bibbona and S. Ditlevsen, Estimation in Discretely Observed Diffusions Killed at a Threshold, Scand J. Stat., 40 (2012), 274-293. |
| [7] | R. Kobayashi and P. Lansky, Estimation of the synaptic input firing rates and characterization of the stimulation effects in an auditory neuron, Front. Comput. Neurosci., 9 (2015), 59. |
| [8] | S. Ditlevsen and A. Samson, Parameter estimation in neuronal stochastic differential equation models from intracellular recordings of membrane potentials in single neurons: a Review, J. de la Socié 157 (2016), 6-16. |
| [9] | U. Picchini, S. Ditlevsen, A. De Gaetano, et al., Parameters of the Diffusion Leaky Integrate-andFire Neuronal Model for a Slowly Fluctuating Signal, Neural Comput., 20 (2008), 2696-2714. |
| [10] | U. Picchini, S. Ditlevsen and A. De Gaetano, Stochastic differential mixed-effects models, Scand J. Stat., 37 (2010), 67-90. |
| [11] | C. Dion, Nonparametric estimation in a mixed-effect Ornstein-Uhlenbeck model, Metrika, 79 (2016), 919-951. |
| [12] | A. N. Burkitt, A review of the integrate-and-fire neuron model: Ⅱ. Inhomogeneous synaptic input and network properties, Biol. Cybern., 95 (2006), 97-112. |
| [13] | W. Gerstner, W.M. Kistler, R. Naud and L. Paninski, Neuronal Dynamics From single neurons to networks and models of cognition, University Press, 2014. |
| [14] | N. Brunel, V. Hakim and M. J. Richardson, Firing-rate resonance in a generalized integrate-andfire neuron with subthreshold resonance, Phys. Rev. E, 67 (2003), 051916. |
| [15] | A. Buonocore, L. Caputo, A. G. Nobile, et al., Gauss-Markov processes in the presence of a reflecting boundary and applications in neuronal models, Appl. Math. Comput., 232 (2014), 799- 809. |
| [16] | G. D'Onofrio and E. Pirozzi, Successive spike times predicted by a stochastic neuronal model with a variable input signal, Math. Biosci. Eng., 13 (2016), 495-507. |
| [17] | A. Buonocore, L. Caputo, A. G. Nobile, et al., Restricted Ornstein Uhlenbeck process and applications in neuronal models with periodic input signals, J. Comput. Appl. Math., 285 (2015), 59-71. |
| [18] | A. R. Bulsara, T. C. Elston, C. R. Doering, et al., Cooperative behavior in periodically driven noisy integrate-fire models of neuronal dynamics, Phys. Rev. E, 53 (1996), 3958-3969. |
| [19] | A. Longtin, Stochastic resonance in neuron models, J. Stat. Phys., 70 (1993), 309-327. |
| [20] | K. Pakdaman, S. Tanabe and T. Shimokawa, Coherence resonance and discharge time reliability in neurons and neuronal models, Neural Netw., 14 (2001), 895-905. |
| [21] | A. Buonocore, L. Caputo and E. Pirozzi, On the evaluation of firing densities for periodically driven neuron models, Math. Biosci., 214 (2008), 122-133. |
| [22] | M. Giraudo, P. Greenwood and L. Sacerdote, How sample paths of leaky integrate-and-fire models are influenced by the presence of a firing threshold, Neural Comput., 23 (2011), 1743-1767. |
| [23] | L. M. Ricciardi, A. Di Crescenzo, V. Giorno, et al., An outline of theoretical and algorithmic approaches to first passage time problems with applications to biological modeling, Math. Japonica, 50 (1999), 247-322. |
| [24] | D. P. Kroese, T. Taimre and Z. I. Botev, Handbook of Monte Carlo Methods, Wiley Series in Probability and Statistics, John Wiley & Sons, Hoboken, NJ, 2011. |
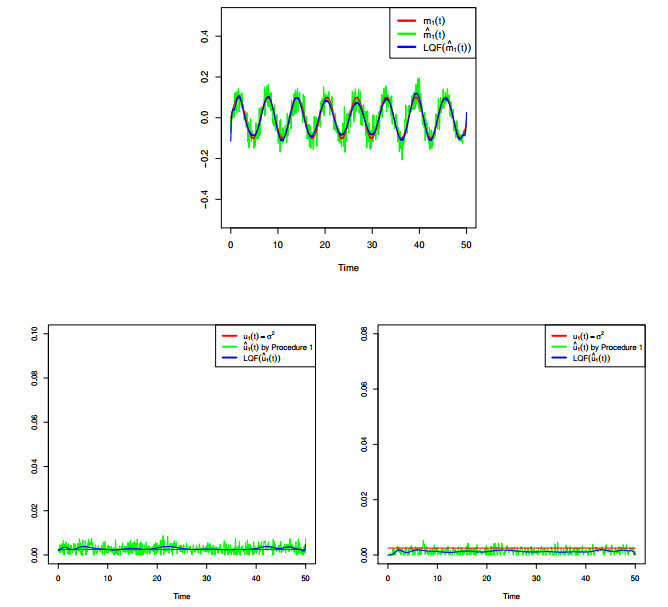
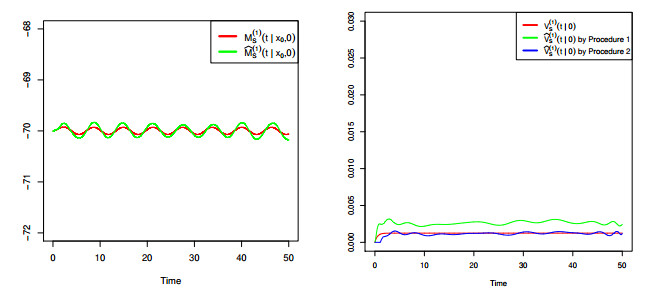
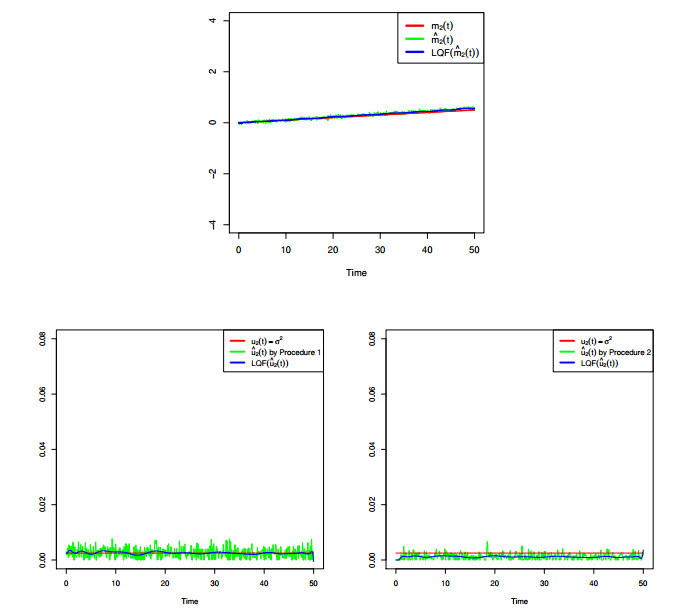
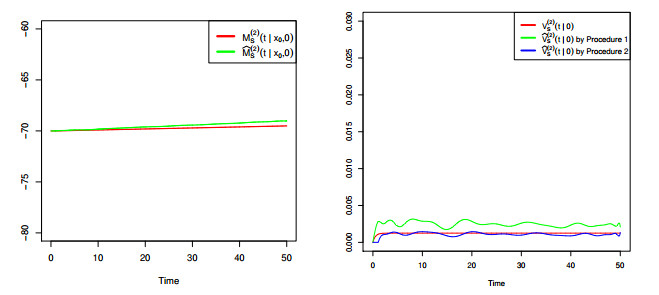
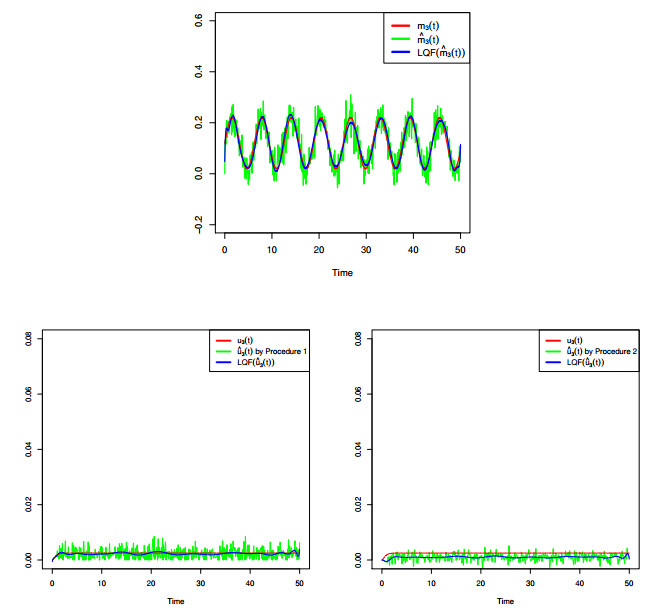
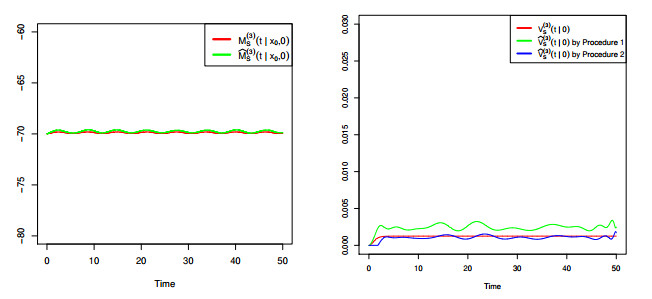
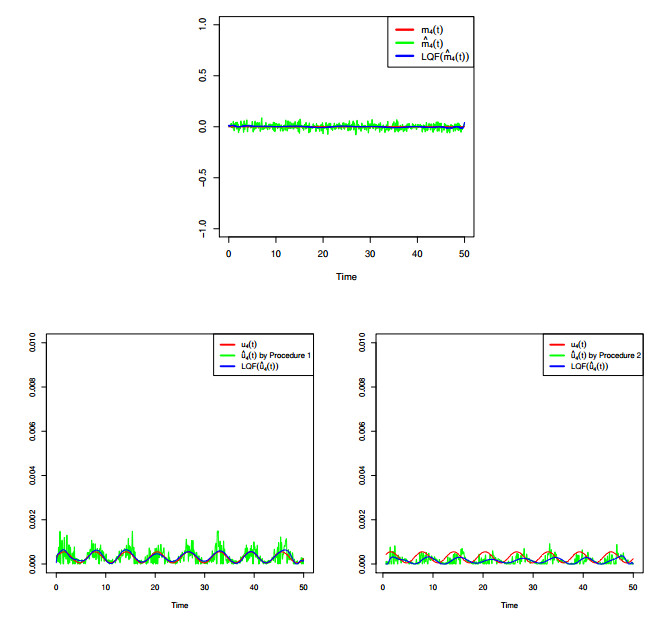
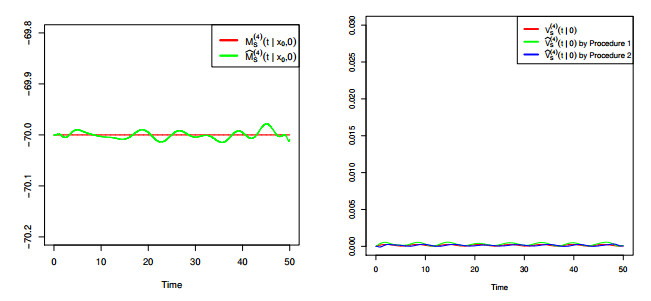
Figures(8) / Tables(4)

Giuseppina Albano, Virginia Giorno. Inference on the effect of non homogeneous inputs in Ornstein-Uhlenbeck neuronal modeling[J]. Mathematical Biosciences and Engineering, 2020, 17(1): 328-348. doi: 10.3934/mbe.2020018







 DownLoad:
DownLoad: