Citation: Maria Pia Saccomani, Karl Thomaseth. Calculating all multiple parameter solutions of ODE models to avoid biological misinterpretations[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 6438-6453. doi: 10.3934/mbe.2019322

| [1] | J. DiStefano, Dynamic System Biology Modeling and Simulation, 1st ed., Academic Press, Elsevier, USA, 2014. |
| [2] | L. Ljung, System Identification - Theory For the User, 2nd ed., PTR, Prentice Hall, Upper Saddle River, N.J., USA, 1999. |
| [3] | E.D. Sontag, Mathematical Control Theory: Deterministic Finite Dimensional Systems, 2nd ed., Springer, New York, 1998. |
| [4] | L. Ljung and S. T. Glad, On global identifiability for arbitrary model parameterizations, Automatica, 30(1994), 265–276. |
| [5] | A. Raue, J. Karlsson, M. P. Saccomani, et al., Comparison of approaches for parameter identifiability analysis of biological systems, Bioinformatics, 30(2014), 1440–1448. |
| [6] | M. P. Saccomani, S. Audoly and L. D'Angiò, Parameter identifiability of nonlinear systems: the role of initial conditions, Automatica, 39(2003), 619–632. |
| [7] | J. D. Stigter and J. Molenaar, A fast algorithm to assess local structural identifiability, Automatica, 58(2015), 118–124. |
| [8] | H. Hong, A. Ovchinnikov, G. Pogudin, et al., SIAN: software for structural identifiability analysis of ODE models, Bioinformatics, (2019), bty1069. |
| [9] | T. S. Ligon, F. Fröhlich, O. T. Chis, et al., GenSSI 2.0: multi-experiment structural identifiability analysis of SBML models, Bioinformatics, 34(2018), 1421–1423. |
| [10] | A. F. Villaverde, A. Barreiro and A. Papachristodoulou, Structural identifiability of dynamic systems biology models, PLoS Comput. Biol., 12(2016), e1005153. |
| [11] | M. Anguelova, J. Karlsson and M. Jirstrand, Minimal output sets for identifiability, Math. Biosci., 239(2012), 139–153. |
| [12] | A. Raue, C. Kreutz, T. Maiwald, et al., Addressing Parameter Identifiability by Model-based Experimentation, IET Syst. Biol., 5(2011), 120–130. |
| [13] | M. Rodriguez-Fernandez, M. Rehberg, A. Kremling, et al., Simultaneous model discrimination and parameter estimation in dynamic models of cellular systems, BMC Syst. Biol., 7(2013), 1–14. |
| [14] | T. R. B. Grandjean, M. J. Chappell, J. W. T. Yates, et al., Structural identifiability analysis of candidate models for in vitro Pitavastatin hepatic uptake, European J. Pharmac. Sci., 46(2013), 259–271. |
| [15] | D. L. I. Janzén, L. Bergenholm, M. Jirstrand, et al., Parameter identifiability of fundamental pharmacodynamic models, Front. Physiol., 7(2016), 590. |
| [16] | H. Klett, M. Rodriguez-Fernandez, S. Dineen, et al., Modeling the inflammatory response in the hypothalamus ensuing heat stroke: Iterative cycle of model calibration, identifiability analysis, experimental design and data collection, Math. Biosci., 260(2015), 35–46. |
| [17] | M. P. Saccomani, An effective automatic procedure for testing parameter identifiability of HIV/AIDS models, B. Math. Biol., 73(2011), 1734–1753. |
| [18] | K. Thomaseth, J. J. Batzel, M. Bachar, et al., Parameter estimation of a model for baroreflex control of unstressed volume, in Mathematical Modeling and Validation in Physiology (eds. J.J. Batzel, M. Bachar, F. Kappel), Springer-Verlag, Berlin, (2013), 215–246. |
| [19] | J. A. Egea, D. Henriques, T. Cokelaer, et al., MEIGO: an open-source software suite based on metaheuristics for global optimization in systems biology and bioinformatics, BMC Bioinformatics, 15(2014), 136. |
| [20] | T. Maiwald and J. Timmer, Dynamical Modeling and Multi-Experiment Fitting with PottersWheel, Bioinformatics, 24(2008), 2037–2043. |
| [21] | G. Bellu, M. P. Saccomani, S. Audoly, et al., DAISY: A new software tool to test global identifiability of biological and physiological systems, Comp. Meth. Prog. Biom., 88(2007), 52–61. |
| [22] | J. F. Ritt, Differential Algebra, Providence, RI: American Mathematical Society, (1950). |
| [23] | S. Audoly, G. Bellu, L. D'Angiò, et al., Global identifiability of nonlinear models of biological systems, IEEE Transact. Biomed. Eng., 48(2001), 55–65. |
| [24] | K. Thomaseth and M. P. Saccomani, Local identifiability analysis of nonlinear ODE models: how to determine all candidate solutions, IFAC PapersOnLine, 51(2018), 529–534. |
| [25] | P. Stapor, D. Weindl, B. Ballnus, et al., PESTO Parameter EStimation TOolbox, Bioinformatics, 34(2008), 705–707. |
| [26] | M. P. Saccomani and K. Thomaseth, Structural vs Practical Identifiability of Nonlinear Differential Equation Models in Systems Biology, in Dynamics of Mathematical Models in Biology (eds. A. Rogato, V. Zazzu, M.R. Guarracino), Springer International Publishing, Switzerland, (2016), 31–42. |
| [27] | J. P. Norton, An Investigation of the Sources of Nonuniqueness in Deterministic Identifiability, Math. Biosci., 60(1982), 89–108. |
| [28] | A. S. Perelson, D. E. Kirschner and R. DeBoer, Dynamics of HIV Infection of CD4+ T cells, Math. Biosci., 114(1993), 81–125. |
| [29] | R. Ferrentino and C. Boniello, On the Well-Posedness for Optimization Problems: A Theoretical Investigation, Appl. Math., 10(2019), 19–38. |
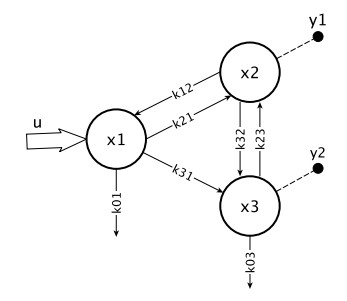
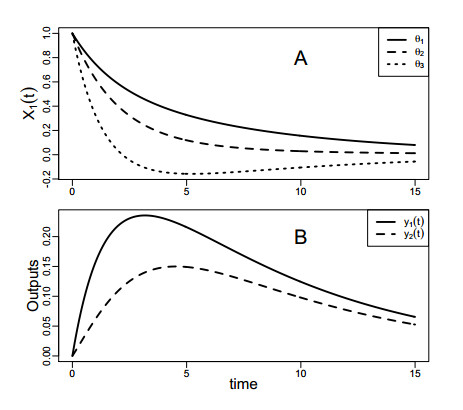
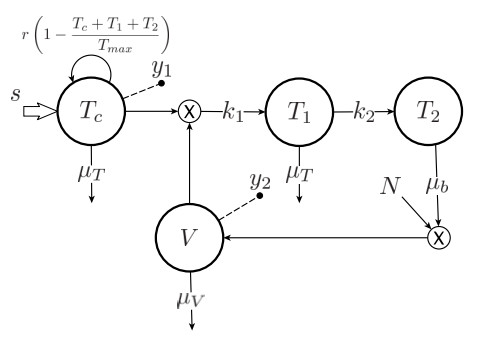
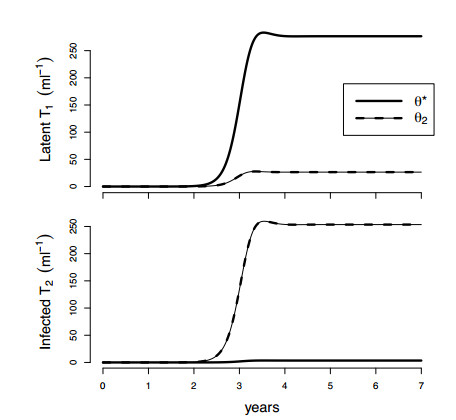
Figures(4) / Tables(2)

Maria Pia Saccomani, Karl Thomaseth. Calculating all multiple parameter solutions of ODE models to avoid biological misinterpretations[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 6438-6453. doi: 10.3934/mbe.2019322







 DownLoad:
DownLoad: