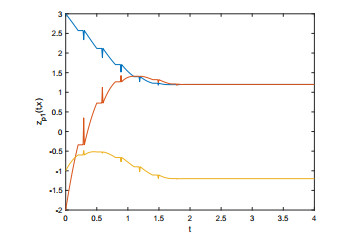
In this paper, we investigate the prespecified-time bipartite synchronization (PTBS) of coupled reaction-diffusion memristive neural networks (CRDMNNs) with both competitive and cooperative interactions. Two types of bipartite synchronization are considered: leaderless PTBS and leader-following PTBS. With the help of a structural balance condition, the criteria for PTBS for CRDMNNs are derived by designing suitable Lyapunov functionals and novel control protocols. Different from the traditional finite-time or fixed-time synchronization, the settling time obtained in this paper is independent of control gains and initial values, which can be pre-set according to the task requirements. Lastly, numerical simulations are given to verify the obtained results.
Citation: Ruoyu Wei, Jinde Cao. Prespecified-time bipartite synchronization of coupled reaction-diffusion memristive neural networks with competitive interactions[J]. Mathematical Biosciences and Engineering, 2022, 19(12): 12814-12832. doi: 10.3934/mbe.2022598
In this paper, we investigate the prespecified-time bipartite synchronization (PTBS) of coupled reaction-diffusion memristive neural networks (CRDMNNs) with both competitive and cooperative interactions. Two types of bipartite synchronization are considered: leaderless PTBS and leader-following PTBS. With the help of a structural balance condition, the criteria for PTBS for CRDMNNs are derived by designing suitable Lyapunov functionals and novel control protocols. Different from the traditional finite-time or fixed-time synchronization, the settling time obtained in this paper is independent of control gains and initial values, which can be pre-set according to the task requirements. Lastly, numerical simulations are given to verify the obtained results.

| [1] |
L. O. Chua, Memristor-the missing circuit element, IEEE Trans. Circuit Theory., 18 (1971), 507–519. https://doi.org/10.1109/TCT.1971.1083337 doi: 10.1109/TCT.1971.1083337

|
| [2] |
D. Strukov, G. Snider, D. Stewart, R. S. Williams, The missing memristor found, Nature, 453 (2008), 80–83. https://doi.org/10.1038/nature06932 doi: 10.1038/nature06932
|
| [3] |
K. Miller, K. S. Nalwa, A. Bergerud, N. M. Neihart, S. Chaudhary, Memristive behavior in thin anodic titania, IEEE Electron Device Lett., 37 (2010), 737–739. https://doi.org/10.1109/LED.2010.2049092 doi: 10.1109/LED.2010.2049092
|
| [4] |
J. Sun, Y. Shen, Q. Yin, C. Xu, Compound synchronization of four memristor chaotic oscillator systems and secure communication, Chaos, 23 (2012), 1–10. https://doi.org/10.1063/1.4794794 doi: 10.1063/1.4794794
|
| [5] |
F. Corinto, A. Ascoli, M. Gilli, Nonlinear dynamics of memristor oscillators, IEEE Trans. Circuits Syst. I, 58 (2011), 1323–1336. https://doi.org/10.1109/TCSI.2010.2097731 doi: 10.1109/TCSI.2010.2097731
|
| [6] |
Y. V. Pershin, M. Di Ventra, Experimental demonstration of associative memory with memristive neural networks, Neural Netw., 23 (2010), 881–886. https://doi.org/10.1016/j.neunet.2010.05.001 doi: 10.1016/j.neunet.2010.05.001
|
| [7] |
L. Wang, H. He, Z. Zeng, Global synchronization of fuzzy memristive neural networks with discrete and distributed delays, IEEE Trans. Fuzz. Syst., 28 (2020), 2022–2034. https://doi.org/10.1109/TFUZZ.2019.2930032 doi: 10.1109/TFUZZ.2019.2930032
|
| [8] |
R. Wei, J. Cao, W. Qian, C. Xue, X. Ding, Finite-time and fixed-time stabilization of inertial memristive Cohen-Grossberg neural networks via non-reduced order method, AIMS Math., 6 (2021), 6915–6932. https://doi.org/10.3934/math.2021405 doi: 10.3934/math.2021405
|
| [9] |
L. Wang, Z. Zeng, M. Ge, A disturbance rejection framework for finite-time and fixed-time stabilization of delayed memristive neural networks, IEEE Trans. Syst. Man Cybern. Syst., 51 (2021), 905–915. https://doi.org/10.1109/TSMC.2018.2888867 doi: 10.1109/TSMC.2018.2888867
|
| [10] |
Y. Sheng, H. Zhang, Z. Zeng, Stability and robust stability of stochastic reaction-diffusion neural networks with infinite discrete and distributed delays, IEEE Trans. Syst. Man. Cybern. Syst., 50 (2020), 1721–1732. https://doi.org/10.1109/TSMC.2017.2783905 doi: 10.1109/TSMC.2017.2783905
|
| [11] |
Y. Sheng, Z. Zeng, Passivity and robust passivity of stochastic reaction-diffusion neural networks with time-varying delays, J. Franklin Inst., 354 (2017), 3995–4012. https://doi.org/10.1016/j.jfranklin.2017.03.014 doi: 10.1016/j.jfranklin.2017.03.014
|
| [12] |
Z. Wang, J. Cao, G. Lu, M. Abdel-Aty, Fixed-time passification analysis of interconnected memristive reaction-diffusion neural networks, IEEE Trans. Netwowk Sci. Eng., 7 (2020), 1814–1824. https://doi.org/10.1109/TNSE.2019.2954463 doi: 10.1109/TNSE.2019.2954463
|
| [13] |
Z. Guo, S. Wang, J. Wang, Global exponential synchronization of coupled delayed memristive neural networks with reaction-diffusion terms via distributed pinning controls, IEEE Trans. Neural Networks Learn. Syst., 32 (2021), 105–116. https://doi.org/10.1109/TNNLS.2020.2977099 doi: 10.1109/TNNLS.2020.2977099
|
| [14] |
X. Yang, J. Cao, Z. Yang, Synchronization of coupled reaction-diffusion neural networks with time-varying delays via pining-impulsive controller, Siam J. Control Optim., 51 (2013), 3486–3510. https://doi.org/10.1137/120897341 doi: 10.1137/120897341
|
| [15] |
C. Altafini, Consensus problems on networks with antagonistic interactions, IEEE Trans. Autom. Control, 58 (2013), 935–946. https://doi.org/10.1109/TAC.2012.2224251 doi: 10.1109/TAC.2012.2224251
|
| [16] |
J. Hu, Y. Wu, T. Li, B. K. Ghosh, Consensus control of general linear multiagent systems with antagonistic interactions and communication noises, IEEE Trans. Autom. Control, 64 (2019), 2122–2127. https://doi.org/10.1109/TAC.2018.2872197 doi: 10.1109/TAC.2018.2872197
|
| [17] |
Y. Wu, L. Liu, J. Hu, G. Feng, Adaptive antisynchronization of multilayer reaction-diffusion neural networks, IEEE Trans. Neural Netw. Learn. Syst., 29 (2018), 807–818. https://doi.org/10.1109/TNNLS.2017.2647811 doi: 10.1109/TNNLS.2017.2647811
|
| [18] |
F. Liu, Q. Song, G. Wen, J. Cao, X. Yang, Bipartite synchronization in coupled delayed neural networks under pinning control, Neural Networks, 108 (2018), 146–154. https://doi.org/10.1016/j.neunet.2018.08.009 doi: 10.1016/j.neunet.2018.08.009
|
| [19] |
N. Li, W. Zheng, Bipartite synchronization of multiple memristor-based neural networks with antagonistic interactions, IEEE Trans. Neural Networks Learn. Syst., 32 (2021), 1642–1653. https://doi.org/10.1109/TNNLS.2020.2985860 doi: 10.1109/TNNLS.2020.2985860
|
| [20] |
N. Li, W. Zheng, Bipartite synchronization for inertia memristor-based neural networks on coopetition networks, Neural Networks, 124 (2020), 39–49. https://doi.org/10.1016/j.neunet.2019.11.010 doi: 10.1016/j.neunet.2019.11.010
|
| [21] |
K. Mao, X. Liu, J. Cao, Y. Hu, Finite-time bipartite synchronization of coupled neural networks with uncertain parameters, Phys. A, 585 (2022), 126431. https://doi.org/10.1016/j.physa.2021.126431 doi: 10.1016/j.physa.2021.126431
|
| [22] |
X. Li, D. Peng, J. Cao, Lyapunov stability for impulsive systems via event-triggered impulsive control, IEEE Trans. Autom. Control, 65 (2020), 4908–4913. https://doi.org/10.1109/TAC.2020.2964558 doi: 10.1109/TAC.2020.2964558
|
| [23] |
X. Li, X. Yang, J. Cao, Event-triggered impulsive control for nonlinear delay systems, Automatica, 117 (2020), 108981. https://doi.org/10.1016/j.automatica.2020.108981 doi: 10.1016/j.automatica.2020.108981
|
| [24] |
S. P. Bhat, D. S. Bernstein, Finite-time stability of continuous autonomous systems, Siam J. Control Optim., 38 (2000), 751–766. https://doi.org/10.1137/S0363012997321358 doi: 10.1137/S0363012997321358
|
| [25] |
X. Li, D. W. C. Ho, J. Cao, Finite-time stability and settling-time estimation of nonlinear impulsive systems, Automatica, 99 (2019), 361–368. https://doi.org/10.1016/j.automatica.2018.10.024 doi: 10.1016/j.automatica.2018.10.024
|
| [26] |
Z. Zhang, X. Liu, D. Zhou, C. Lin, J. Chen, H. Wang, Finite-time stabilizability and instabilizability for complex-valued memristive neural networks with time delays, IEEE Trans. Syst. Man Cybern. Syst., 48 (2018), 2371–2382. https://doi.org/10.1109/TSMC.2017.2754508 doi: 10.1109/TSMC.2017.2754508
|
| [27] |
L. Feng, J. Yu, C. Hu, C. Yang, H. Jiang, Nonseparation method-based finite/fixed-time synchronization of fully complex-valued discontinuous neural networks, IEEE Trans. Cybern., 51 (2021), 3212–3223. https://doi.org/10.1109/TCYB.2020.2980684 doi: 10.1109/TCYB.2020.2980684
|
| [28] |
A. Polyakov, Nonlinear feedback design for fixed-time stabilization of linear control systems, IEEE Trans. Automat. Control, 57 (2012), 2106–2110. https://doi.org/10.1109/TAC.2011.2179869 doi: 10.1109/TAC.2011.2179869
|
| [29] |
R. Wei, J. Cao, Fixed-time synchronization of second-order MNNs in quaternion field, IEEE Trans. Syst. Man Cybern. Syst., 51 (2021), 3587–3598. https://doi.org/10.1109/TSMC.2019.2931091 doi: 10.1109/TSMC.2019.2931091
|
| [30] |
C. Hu, H. Jiang, Special functions-based fixed-time estimation and stabilization for dynamic systems, IEEE Trans. Syst. Man Cybern. Syst., 52 (2022), 3251–3262. https://doi.org/10.1109/TSMC.2021.3062206 doi: 10.1109/TSMC.2021.3062206
|
| [31] |
X. Ding, J. Cao, A. Alsaedi, T. Hayat, Robust fixed-time synchronization for uncertain complex-valued neural networks with discontinuous activation functions, Neural Networks, 90 (2017), 42–55. https://doi.org/10.1016/j.neunet.2017.03.006 doi: 10.1016/j.neunet.2017.03.006
|
| [32] |
C. Hu, H. He, H. Jiang, Fixed/Preassigned-time synchronization of quaternion-valued neural networks via pure power-law control, Neural Networks, 146 (2022), 341–349. https://doi.org/10.1016/j.neunet.2021.11.023 doi: 10.1016/j.neunet.2021.11.023
|
| [33] |
C. Hu, H. He, H. Jiang, Fixed/preassigned-time synchronization of complex networks via improving fixed-Time stability, IEEE Trans. Cybern., 51 (2021), 2882–2892. https://doi.org/10.1109/TCYB.2020.2977934 doi: 10.1109/TCYB.2020.2977934
|
| [34] |
S. Shao, X. Liu, J. Cao, Prespecified-time synchronization of switched coupled neural networks via smooth controllers, Neural Networks, 133 (2021), 32–39. https://doi.org/10.1016/j.neunet.2020.10.007 doi: 10.1016/j.neunet.2020.10.007
|
| [35] |
X. Liu, D. W. C. Ho, and C. Xie, Prespecified-time cluster synchronization of complex networks via a smooth control approach, IEEE Trans. Cybern., 50 (2020), 1771–1775. https://doi.org/10.1109/TCYB.2018.2882519 doi: 10.1109/TCYB.2018.2882519
|


Figures(3)

Ruoyu Wei, Jinde Cao. Prespecified-time bipartite synchronization of coupled reaction-diffusion memristive neural networks with competitive interactions[J]. Mathematical Biosciences and Engineering, 2022, 19(12): 12814-12832. doi: 10.3934/mbe.2022598







 DownLoad:
DownLoad: