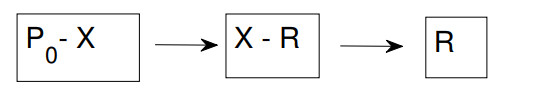
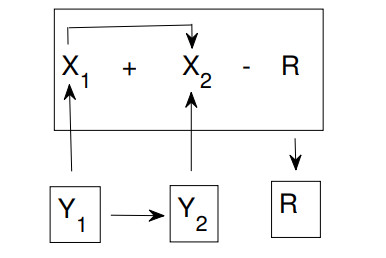
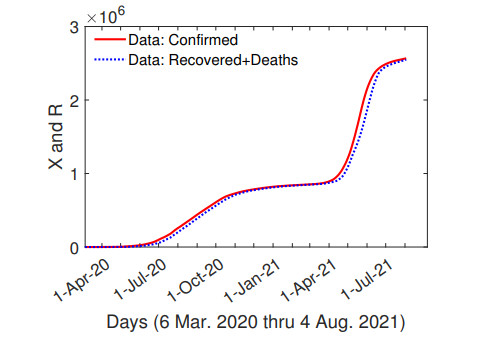
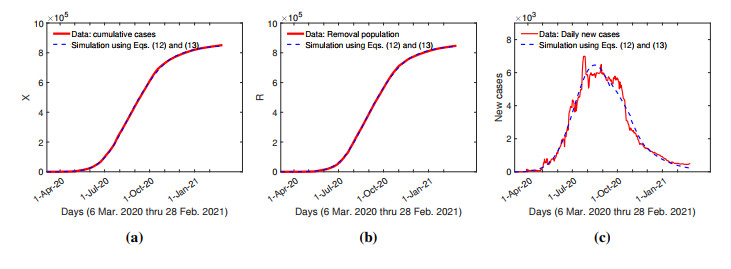
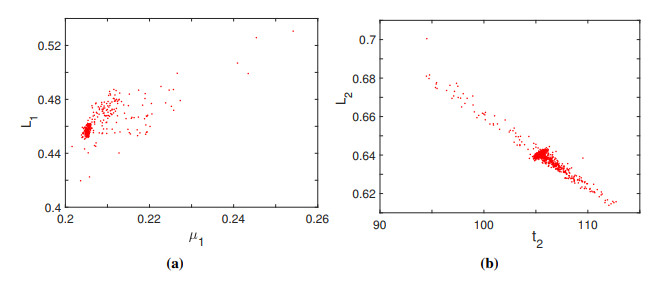
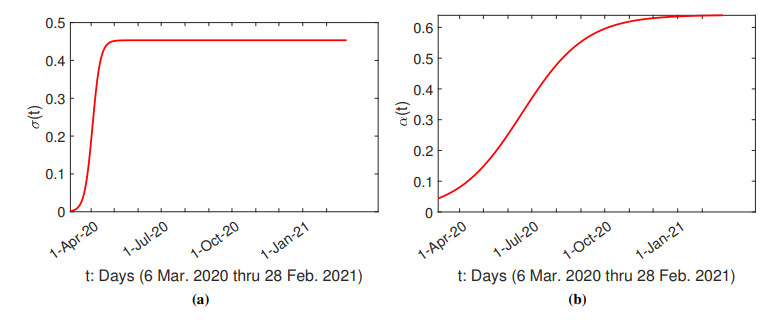
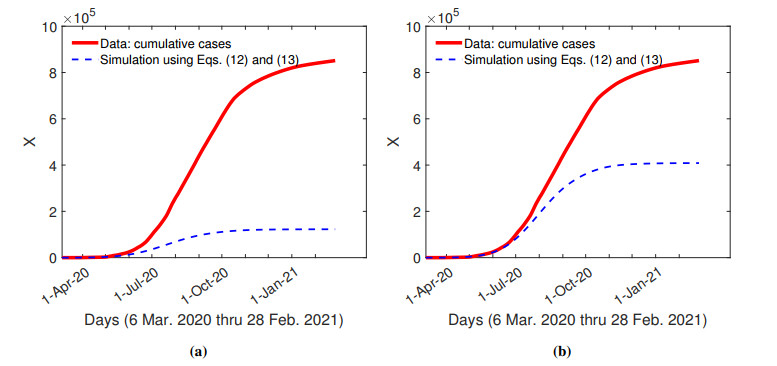
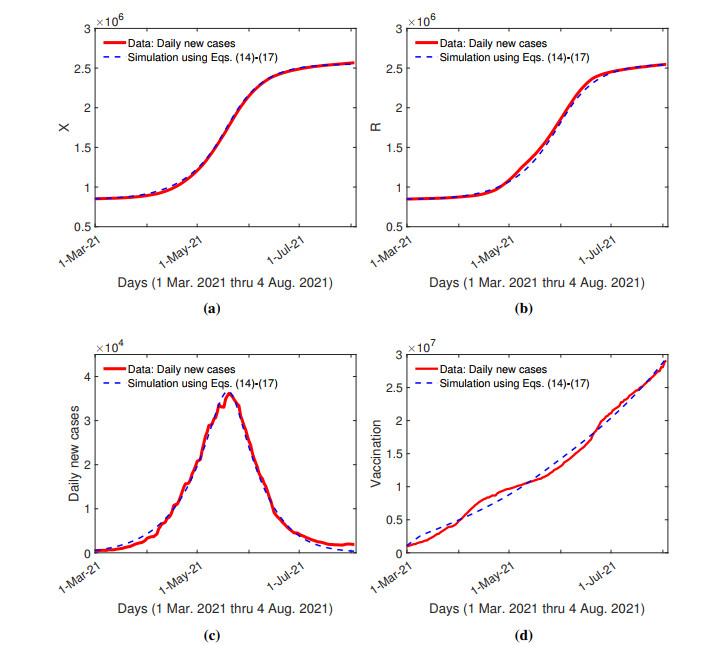
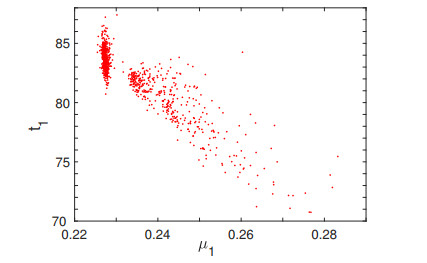
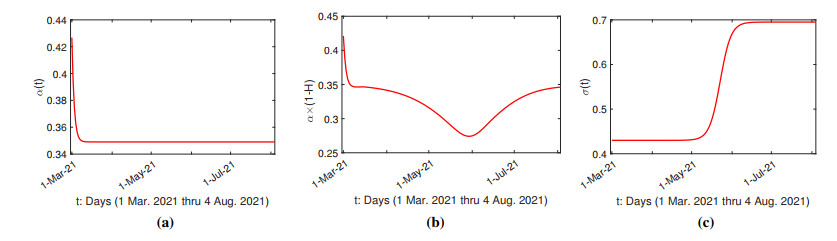
The purpose of this paper is to build a mathematical model for the study of the roles of lock-down, social distancing, vaccination, detection efficiency, and health care capacity planning of the COVID-19 pandemic taking into account the demographic topology of the State of Tamil Nadu, India. Two mathematical models are proposed for the evolution of the first and second wave of COVID-19 pandemic. The model for the first wave considers lock-down orders, social distancing measures, and detection efficiency. The model for the second wave considers more sub-populations and incorporates two more elements, vaccination and health care capacity. Daily reported data on the evolution of the COVID-19 pandemic are used to determine the parameter values. The dynamics produced by the mathematical model closely follow the evolution of COVID-19 in the State of Tamil Nadu. Numerical simulation shows that the lock-down effect is limited. Social distancing implementation and detection of positive cases are relatively ineffective compared with other big cities. Shortage of health care resources is one of the factors responsible for rapidly spreading in the second wave in Tamil Nadu.
Citation: Avinash Shankaranarayanan, Hsiu-Chuan Wei. Mathematical modeling of SARS-nCoV-2 virus in Tamil Nadu, South India[J]. Mathematical Biosciences and Engineering, 2022, 19(11): 11324-11344. doi: 10.3934/mbe.2022527
The purpose of this paper is to build a mathematical model for the study of the roles of lock-down, social distancing, vaccination, detection efficiency, and health care capacity planning of the COVID-19 pandemic taking into account the demographic topology of the State of Tamil Nadu, India. Two mathematical models are proposed for the evolution of the first and second wave of COVID-19 pandemic. The model for the first wave considers lock-down orders, social distancing measures, and detection efficiency. The model for the second wave considers more sub-populations and incorporates two more elements, vaccination and health care capacity. Daily reported data on the evolution of the COVID-19 pandemic are used to determine the parameter values. The dynamics produced by the mathematical model closely follow the evolution of COVID-19 in the State of Tamil Nadu. Numerical simulation shows that the lock-down effect is limited. Social distancing implementation and detection of positive cases are relatively ineffective compared with other big cities. Shortage of health care resources is one of the factors responsible for rapidly spreading in the second wave in Tamil Nadu.

| [1] | World Health Organization, Naming the coronavirus disease (COVID-19) and the virus that causes it. Available from: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/technical-guidance/naming-the-coronavirus-disease-(covid-2019)-and-the-virus-that-causes-it. |
| [2] |
Coronaviridae Study Group of the International Committee on Taxonomy of Viruses, The species Severe acute respiratory syndrome-related coronavirus: classifying 2019-nCoV and naming it SARS-CoV-2, Nat. Microbiol., 5 (2020), 536–544. https://doi.org/10.1038/s41564-020-0695-z doi: 10.1038/s41564-020-0695-z

|
| [3] |
M. Mohammed, H. Syamsudin, S. Al-Zubaidi, A. Sairah, R. Ramli, E. Yusuf, Novel COVID-19 detection and diagnosis system using IOT based smart helmet, Int. J. Psychosoc. Rehabilitation, 24 (2020), 2296–2303. https://doi.org/10.37200/IJPR/V24I7/PR270221 doi: 10.37200/IJPR/V24I7/PR270221
|
| [4] | World Health Organization, WHO Director-General's opening remarks at the media briefing on COVID-19-11 March 2020. Available from: https://www.who.int/dg/speeches/detail/who-director-general-s-openingremarks-at-the-media-briefing-on-covid-19—11- march-2020. |
| [5] |
C. Huang, Y. Wang, X. Li, L. Ren, J. Zhao, Y. Hu, et al, Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China, The lancet, 395 (2020), 497–506. https://doi.org/10.1016/S0140-6736(20)30183-5 doi: 10.1016/S0140-6736(20)30183-5
|
| [6] |
H. Lu, C. W. Stratton, Y. W. Tang, Outbreak of pneumonia of unknown etiology in Wuhan, China: The mystery and the miracle, J. Med. Virol., 92 (2020), 401–402. https://doi.org/10.1002/jmv.25678 doi: 10.1002/jmv.25678
|
| [7] | World Health Organization, management of severe acute respiratory infections when novel coronavirus is suspected: what to do and what not to do, Available from: https://www.who.int/csr/disease/coronavirus_infections/InterimGuidance_ClinicalManagem- ent_NovelCoronavirus. |
| [8] |
S. Kashte, A. Gulbake, S. El-Amin, A. Gupta, COVID-19 vaccines: rapid development, implications, challenges and future prospects, Human. Cell, 34 (2021), 1–23. https://doi.org/10.1007/s13577-021-00512-4 doi: 10.1007/s13577-021-00512-4
|
| [9] |
T. Balasubramaniam, D. J. Warne, R. Nayak, K. Mengersen, Explainability of the COVID-19 epidemiological model with nonnegative tensor factorization, Int J Data Sci. Anal., 30 (2022), 1–14. https://doi.org/10.1007/s41060-022-00324-1 doi: 10.1007/s41060-022-00324-1
|
| [10] |
R. S. Yadav, Mathematical modeling and simulation of SIR model for COVID-2019 epidemic outbreak: A case study of India, INFOCOMP J. Comput. Sci., 19 (2020), 1–9. https://doi.org/10.1101/2020.05.15.20103077 doi: 10.1101/2020.05.15.20103077
|
| [11] | H. Hassen, A. Elaoud, N. Salah, A. Masmoudi, A SIR-Poisson model for COVID-19: evolution and transmission inference in the Maghreb central regions, Arab. J. Sci. Eng.. 46 (2021), 93–102. https://doi.org/10.1007/s13369-020-04792-0 |
| [12] |
R. Bhardwaj, A predictive model for the evolution of COVID-19, Trans. Indian Natl. Acad. Eng., 5 (2020), 133–140. https://doi.org/10.1007/s41403-020-00130-w doi: 10.1007/s41403-020-00130-w
|
| [13] |
B. Jamshidi, M. Rezaei, S. J. Zargaran, F. Najafi, Mathematical modeling the epicenters of coronavirus disease-2019 (COVID-19) pandemic, Epidemiol. Methods, 9 (2020), 20200009. https://doi.org/10.1515/em-2020-0009 doi: 10.1515/em-2020-0009
|
| [14] |
K. Santosh, COVID-19 prediction models and unexploited data, J. Med. Syst., 44 (2021), 170. https://doi.org/10.1007/s10916-020-01645-z doi: 10.1007/s10916-020-01645-z
|
| [15] |
A. L. Jenner, A. A. Rosemary, S. Alfonso, V. Crowe, X. Deng, A. P. Smith, et al., COVID-19 virtual patient cohort suggests immune mechanisms driving disease outcomes, Plos Pathog., 17 (2021), e1009753. https://doi.org/10.1371/journal.ppat.1009753 doi: 10.1371/journal.ppat.1009753
|
| [16] |
S. Farhang-Sardroodi, C. S. Korosec, S. Gholami, M. Craig, I. R. Moyles, M. S. Ghaemi, et al., Analysis of hostimmunological response of Adenovirus-based COVID-19 vaccines, Vaccines, 9 (2021), 861. https://doi.org/10.3390/vaccines9080861 doi: 10.3390/vaccines9080861
|
| [17] | A. Goyal, E. F. Cardozo-Ojeda, J. T. Schiffer, Potency and timing of antiviral therapy as determinants of duration of SARS-CoV-2 shedding and intensity of inflammatory response, Sci. Adv., 6 (2020), eabc7112. https://www.science.org/doi/10.1126/sciadv.abc7112 |
| [18] |
M. Dawoudi, Mathematical modeling approaches to understanding severe acute respiratory syndrome coronavirus 2 (SARSCoV-2) DNA sequences linked coronavirus disease (COVID-19) for discovery of potential new drugs, OAJBS, 2 (2020), 316–317. https://doi.org/10.38125/OAJBS.000173 doi: 10.38125/OAJBS.000173
|
| [19] | M. Wanjau, Mathematical modeling of COVID-19 transmission with mass testing and contact tracing, J. Math., 16 (2020), 55–64. |
| [20] |
J. Ndam, Modelling the impacts of lockdown and isolation on the eradication of COVI-19, Biomath, 9 (2020), 2009107. http://dx.doi.org/10.11145/j.biomath.2020.09.107 doi: 10.11145/j.biomath.2020.09.107
|
| [21] |
R. Asempapa, B. Oduro, O. Apenteng, V. Magagula, A COVID-19 mathematical model of at-risk populations with non-pharmaceutical preventive measures: The case of Brazil and South Africa, Infect. Dis. Model., 7 (2022), 45–61. https://doi.org/10.1016/j.idm.2021.11.005 doi: 10.1016/j.idm.2021.11.005
|
| [22] |
J. Rojas-Vallejos, Strengths and limitations of mathematical models in pandemics-the case of COVID-19 in Chile, Medwave, 20 (2020), e7874. https://doi.org/10.5867/medwave.2020.03.7874 doi: 10.5867/medwave.2020.03.7874
|
| [23] |
M. Alvarez, González-González E, Santiago G, Modeling COVID-19 epidemics in an Excel spreadsheet to enable first-hand accurate predictions of the pandemic evolution in urban areas, Sci. Rep., 11 (2021), 1–12. https://doi.org/10.1038/s41598-021-83697-w doi: 10.1038/s41598-021-83697-w
|
| [24] | S. Baharom, S. Anuar, N. Zolkifly, H. Tahir, The people's behavior change during pandemic of Covid-19; the four aspects of design thinking, in International Conference of Innovation in Media and Visual Design, 502 (2020), 180–186. https://doi.org/10.2991/assehr.k.201202.073 |
| [25] |
W. Wolff, C. Martarelli, J. Schüler, M. Bieleke, High boredom proneness and low trait self-control impair adherence to social distancing guidelines during the COVID-19 pandemic, Int. J. Environ. Res. Public Health, 17 (2020), 5420. https://doi.org/10.3390/ijerph17155420 doi: 10.3390/ijerph17155420
|
| [26] |
T. Zhao, K. Xuan, C. Sun, Y. Sun, The importance of social distancing policy, J. Public Health, 43 (2021), e269–e269. https://doi.org/10.1093/pubmed/fdaa219 doi: 10.1093/pubmed/fdaa219
|
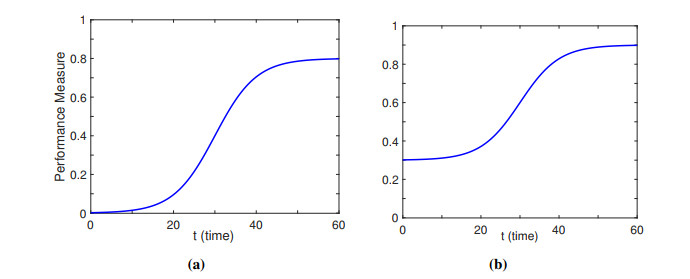
| [27] | J. Murre, S-shaped learning curves, Psychon. Bull. Rev., 21 (2014), 344–356. https://doi.org/10.3758/s13423-013-0522-0 |
| [28] |
T. Netland, K. Ferdows, The S-curve effect of lean implementation, Prod. Oper. Manag., 25 (2016), 1106–1120. https://doi.org/10.1111/poms.12539 doi: 10.1111/poms.12539
|
| [29] | S. Kaushal, A. Rajput, S. Bhattacharya, M. Vidyasagar, A. Kumar, M. Prakash, et al., Estimating the herd immunity threshold by accounting for the hidden asymptomatics using a COVID-19 specific model. Plos One, 15 (2020), e0242132. https://doi.org/10.1371/journal.pone.0242132 |
| [30] | S. SeyedAlinaghi, L. Abbasian, M. Solduzian, N. A. Yazdi, F. Jafari, A. Adibimehr, et al., Predictors of the prolonged recovery period in COVID-19 patients: a cross-sectional study, Eur. J. Med. Res., 26 (2021). https://doi.org/10.1186/s40001-021-00513-x |
| [31] |
J. Lei, M. Li, X. Wang, Predicting the development trend of the second wave of COVID-19 in five European countries, J. Med. Virol., 93 (2021), 5896–5907. https://doi.org/10.1002/jmv.27143 doi: 10.1002/jmv.27143
|
| [32] |
M. Linden, J. Dehning, S. Mohr, J. Mohring, M. Meyer-Hermann, I. Pigeot, Case numbers beyond contact tracing capacity are endangering the containment of COVID-19, Dtsch. Arztebl. Int., 117 (2020), 790–791. https://doi.org/10.3238/arztebl.2020.0790 doi: 10.3238/arztebl.2020.0790
|
| [33] |
E. Argulian, Anticipating the "second wave" of health care strain in the covid-19 pandemic, J. Am. Coll. Cardiol. Case Rep., 2 (2020), 845–846. https://doi.org/10.1016/j.jaccas.2020.04.005 doi: 10.1016/j.jaccas.2020.04.005
|
| [34] |
S. Vaid, A. McAdie, R. Kremer, V. Khanduja, M. Bhandari, Risk of a second wave of Covid-19 infections: using artificial intelligence to investigate stringency of physical distancing policies in North America, Int. Orthop., 44 (2020), 1581–1589. https://doi.org/10.1007/s00264-020-04653-3 doi: 10.1007/s00264-020-04653-3
|
| [35] |
K. R. Nehal, L. M. Steendam, M. C. Ponce, M. van der Hoeven, G. S. A. Smit, Worldwide vaccination willingness for COVID-19: a systematic review and meta-analysis, Vaccines, 9 (2021), 1071. https://doi.org/10.3390/vaccines9101071 doi: 10.3390/vaccines9101071
|
| [36] |
C. Lin, P. Tu, L. Beitsch, Confidence and receptivity for COVID-19 vaccines: a rapid systematic review, Vaccines, 9 (2021), 16. https://doi.org/10.3390/vaccines9010016 doi: 10.3390/vaccines9010016
|
| [37] | Q. Wang, L. Yang, H. Jin, L. Lin, Vaccination against COVID-19: A systematic review and meta-analysis of acceptability and its predictors, Prev. Med., (2021), 2021106694. https://doi.org/10.1016/j.ypmed.2021.106694 |
| [38] | JHU CSSE COVID-19 daily reports, accessed on 8 November 2021. Available from: https://github.com/CSSEGISandData/COVID-19/tree/master/csse_covid_19_data/csse_covid_19_ daily_reports |
| [39] | Daily vaccination reports maintained by our world in data, accessed on 14 December 2021. Available from: https://raw.githubusercontent.com/owid/covid-19-data/master/public/data/vaccinations/vaccinations.csv |
| [40] |
K. Parvathy, Lifestyle as risk factor for breast cancer: a case control study in Chennai, Tamil Nadu, Inida, Int. J. Biol. Sci., 12 (2021), 13–32. https://doi.org/10.53390/ijbs.v12.i1.3 doi: 10.53390/ijbs.v12.i1.3
|
| [41] |
J. Nelder, R. Mead, A simplex method for function minimization, Comput. J., 7 (1965), 308–313. https://doi.org/10.1093/COMJNL/7.4.308 doi: 10.1093/COMJNL/7.4.308
|
| [42] |
T. Luzyanina, S. Mrusek, J. T. Edwards, D. Roose, S. Ehl, G. Bocharov, et al., Computational analysis of CFSE proliferation assay, J. Math. Biol., 54 (2007), 57–89. https://doi.org/10.1007/s00285-006-0046-6 doi: 10.1007/s00285-006-0046-6
|
| [43] |
C. Chakraborty, A. Sharma, M. Bhattacharya, G. Agoramoorthy, S. Lee, The current second wave and COVID-19 vaccination status in India, Brain Behav. Immun., 96 (2021), 1–4. https://doi.org/10.1016/j.bbi.2021.05.018 doi: 10.1016/j.bbi.2021.05.018
|
| [44] |
C. Cai, Y. Peng, E. Shen, Q. Huang, Y. Chen, P. Liu, et al., A comprehensive analysis of the efficacy and safety of COVID-19 vaccines, Mol. Ther., 29 (2021), 2794–2805. https://doi.org/10.1016/j.ymthe.2021.08.001 doi: 10.1016/j.ymthe.2021.08.001
|
| [45] |
F. Polack, S. Thomas, N. Kitchin, J. Absalon, A. Gurtman, S. Lockhart, J. Perez, Safety and efficacy of the BNT162b2 mRNA Covid-19 vaccine, N. Engl. J. Med., 383 (2020), 2603–2615. https://doi.org/10.1056/NEJMoa2034577 doi: 10.1056/NEJMoa2034577
|
| [46] |
V. Jain, K. Iyengar, R. Vaishya, Differences between First wave and Second wave of COVID-19 in India, Diabetes Metab. Syndr., 15 (2021), 1047–1048. https://doi.org/10.1016/j.dsx.2021.05.009 doi: 10.1016/j.dsx.2021.05.009
|
Figures(11) / Tables(4)

Avinash Shankaranarayanan, Hsiu-Chuan Wei. Mathematical modeling of SARS-nCoV-2 virus in Tamil Nadu, South India[J]. Mathematical Biosciences and Engineering, 2022, 19(11): 11324-11344. doi: 10.3934/mbe.2022527







 DownLoad:
DownLoad: