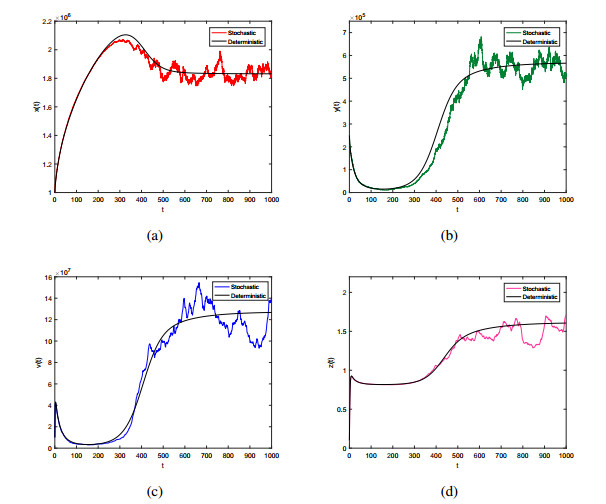
Hepatitis B is a disease that damages the liver, and its control has become a public health problem that needs to be solved urgently. In this paper, we investigate analytically and numerically the dynamics of a new stochastic HBV infection model with antiviral therapies and immune response represented by CTL cells. Through using the theory of stochastic differential equations, constructing appropriate Lyapunov functions and applying Itô's formula, we prove that the disease-free equilibrium of the stochastic HBV model is stochastically asymptotically stable in the large, which reveals that the HBV infection will be eradicated with probability one. Moreover, the asymptotic behavior of globally positive solution of the stochastic model near the endemic equilibrium of the corresponding deterministic HBV model is studied. By using the Milstein's method, we provide the numerical simulations to support the analysis results, which shows that sufficiently small noise will not change the dynamic behavior, while large noise can induce the disappearance of the infection. In addition, the effect of inhibiting virus production is more significant than that of blocking new infection to some extent, and the combination of two treatment methods may be the better way to reduce HBV infection and the concentration of free virus.
Citation: Yiping Tan, Yongli Cai, Zhihang Peng, Kaifa Wang, Ruoxia Yao, Weiming Wang. Dynamics of a stochastic HBV infection model with drug therapy and immune response[J]. Mathematical Biosciences and Engineering, 2022, 19(8): 7570-7585. doi: 10.3934/mbe.2022356
Hepatitis B is a disease that damages the liver, and its control has become a public health problem that needs to be solved urgently. In this paper, we investigate analytically and numerically the dynamics of a new stochastic HBV infection model with antiviral therapies and immune response represented by CTL cells. Through using the theory of stochastic differential equations, constructing appropriate Lyapunov functions and applying Itô's formula, we prove that the disease-free equilibrium of the stochastic HBV model is stochastically asymptotically stable in the large, which reveals that the HBV infection will be eradicated with probability one. Moreover, the asymptotic behavior of globally positive solution of the stochastic model near the endemic equilibrium of the corresponding deterministic HBV model is studied. By using the Milstein's method, we provide the numerical simulations to support the analysis results, which shows that sufficiently small noise will not change the dynamic behavior, while large noise can induce the disappearance of the infection. In addition, the effect of inhibiting virus production is more significant than that of blocking new infection to some extent, and the combination of two treatment methods may be the better way to reduce HBV infection and the concentration of free virus.

| [1] |
D. Anwarud, Y. Li, Stochastic optimal analysis for the hepatitis B epidemic model with Markovian switching, Math. Methods Appl. Sci., 3 (2022), 1–26. https://doi.org/10.1002/mma.8218 doi: 10.1002/mma.8218

|
| [2] |
A. Din, Y. Li, Q. Liu, Viral dynamics and control of hepatitis B virus (HBV) using an epidemic model, Alexandria Eng. J., 59 (2020), 667–679. https://doi.org/10.1016/j.aej.2020.01.034 doi: 10.1016/j.aej.2020.01.034
|
| [3] |
A. Din, Y. Li, Stationary distribution extinction and optimal control for the stochastic hepatitis B epidemic model with partial immunity, Phys. Scr., 96 (2021), 074005. https://doi.org/10.1088/1402-4896/abfacc doi: 10.1088/1402-4896/abfacc
|
| [4] |
A. Din, Y. Li, Lévy noise impact on a stochastic hepatitis B epidemic model under real statistical data and its fractal-fractional Atangana-Baleanu order model, Phys. Scr., 96 (2021), 124008. https://doi.org/10.1088/1402-4896/ac1c1a doi: 10.1088/1402-4896/ac1c1a
|
| [5] |
A. Din, Y. Li, M. A. Shah, The complex dynamics of hepatitis B infected individuals with optimal control, J. Syst. Sci. Complexity, 34 (2021), 1301–1323. https://doi.org/10.1007/s11424-020-0053-5 doi: 10.1007/s11424-020-0053-5
|
| [6] |
J. Danane, K. Allali, Z. Hammouch, Mathematical analysis of a fractional differential model of HBV infection with antibody immune response, Chaos, Solitons Fractals, 136 (2020), 109787. https://doi.org/10.1016/j.chaos.2020.109787 doi: 10.1016/j.chaos.2020.109787
|
| [7] | World Health Organization (WHO), Hepatitis B, Accessed 27 July 2021. Available from: https://www.who.int/news-room/fact-sheets/detail/hepatitis-b. |
| [8] |
D. Grimm, R. Thimme, H. E. Blum, HBV life cycle and novel drug targets, Hepatol. Int., 5 (2011), 644–653. https://doi.org/10.1007/s12072-011-9261-3 doi: 10.1007/s12072-011-9261-3
|
| [9] |
M. A. Nowak, S. Bonhoeffer, A. M. Hill, R. Boehme, H. C. Thomas, H. McDade, Viral dynamics in hepatitis B virus infection, Proc. Natl. Acad. Sci., 93 (1996), 4398–4402. https://doi:10.1073/pnas.93.9.4398 doi: 10.1073/pnas.93.9.4398
|
| [10] |
H. L. Smith, P. D. Leenheer, Virus dynamics: a global analysis, SIAM J. Appl. Math., 63 (2003), 1313–1327. https://doi.org/10.1137/S0036139902406905 doi: 10.1137/S0036139902406905
|
| [11] |
Y. Yang, Y. Xiao, Threshold dynamics for an HIV model in periodic environments, J. Math. Anal. Appl., 361 (2010), 59–68. https://doi.org/10.1016/j.jmaa.2009.09.012 doi: 10.1016/j.jmaa.2009.09.012
|
| [12] |
H. Dahari, E. Shudo, R. Ribeiro, A. Perelson, Modeling complex decay profiles of hepatitis B virus during antiviral therapy, Hepatology, 49 (2009), 32–38. https://doi.org/10.1002/hep.22586 doi: 10.1002/hep.22586
|
| [13] |
T. C. Reluga, H. Dahari, A. S. Perelson, Analysis of hepatitis C virus infection models with hepatocyte homeostasis, SIAM J. Appl. Math., 69 (2009), 999–1023. https://doi.org/10.1137/080714579 doi: 10.1137/080714579
|
| [14] |
M. A. Nowak, C. R. Bangham, Population dynamics of immune responses to persistent viruses, Science, 272 (1996), 74–79. https://doi.org/10.1126/science.272.5258.74 doi: 10.1126/science.272.5258.74
|
| [15] |
M. Tsiang, J. F. Rooney, J. J. Toole, C. S. Gibbs, Biphasic clearance kinetics of hepatitis B virus from patients during adefovir dipivoxil therapy, Hepatology, 29 (1999), 1863–1869. https://doi.org/10.1002/hep.510290626 doi: 10.1002/hep.510290626
|
| [16] |
K. Hattaf, N. Yousfi, A. Tridane, Mathematical analysis of a virus dynamics model with general incidence rate and cure rate, Nonlinear Anal. Real World Appl., 13 (2012), 1866–1872. https://doi.org/10.1016/j.nonrwa.2011.12.015 doi: 10.1016/j.nonrwa.2011.12.015
|
| [17] |
T. Zhang, X. Meng, T. Zhang, Global dynamics of a virus dynamical model with cell-to-cell transmission and cure rate, Comput. Math. Methods Med., 2015 (2015), 1–8. https://doi.org/10.1155/2015/758362 doi: 10.1155/2015/758362
|
| [18] |
S. M. Salman, A. M. Yousef, On a fractional-order model for HBV infection with cure of infected cells, J. Egypt. Math. Soc., 25 (2017), 445–451. https://doi.org/10.1016/j.joems.2017.06.003 doi: 10.1016/j.joems.2017.06.003
|
| [19] |
S. R. Lewin, R. Ribeiro, T. Walters, G. K. Lau, S. Bowden, S. Locarnini, et al., Analysis of hepatitis B viral load decline under potent therapy: complex decay profiles observed, Hepatology, 34 (2001), 1012–1020. https://doi.org/10.1053/jhep.2001.28509 doi: 10.1053/jhep.2001.28509
|
| [20] | K. Hattaf, N. Yousfi, A. Tridane, Global stability analysis of a generalized virus dynamics model with the immune response, Can. Appl. Math. Q., 20 (2012), 499–518. |
| [21] |
X. Wang, Y. Tao, X. Song, Global stability of a virus dynamics model with Beddington-DeAngelis incidence rate and CTL immune response, Nonlinear Dyn., 66 (2011), 825–830. https://doi.org/10.1007/s11071-011-9954-0 doi: 10.1007/s11071-011-9954-0
|
| [22] |
X. Yang, Y. Su, L. Yang, X. Zhuo, Global analysis and simulation of a fractional order HBV immune model, Chaos Solitons Fractals, 154 (2022), 111648. https://doi.org/10.1016/j.chaos.2021.111648 doi: 10.1016/j.chaos.2021.111648
|
| [23] |
P. Yosyingyong, R. Viriyapong, Global stability and optimal control for a hepatitis B virus infection model with immune response and drug therapy, J. Appl. Math. Comput., 60 (2019), 537–565. https://doi.org/10.1007/s12190-018-01226-x doi: 10.1007/s12190-018-01226-x
|
| [24] |
X. Wang, Y. Tan, Y. Cai, K. Wang, W. Wang, Dynamics of a stochastic HBV infection model with cell-to-cell transmission and immune response, Math. Biosci. Eng., 18 (2020), 616–642. https://doi.org/10.3934/mbe.2021034 doi: 10.3934/mbe.2021034
|
| [25] |
Y. Ma, X. Yu, The effect of environmental noise on threshold dynamics for a stochastic viral infection model with two modes of transmission and immune impairment, Chaos Solitons Fractals, 134 (2020), 109699. https://doi.org/10.1016/j.chaos.2020.109699 doi: 10.1016/j.chaos.2020.109699
|
| [26] |
F. A. Rihan, H. J. Alsakaji, C. Rajivganthi, Stochastic SIRC epidemic model with time-delay for COVID-19, Adv. Differ. Equations, 2020 (2020), 1–20. https://doi.org/10.1186/s13662-020-02964-8 doi: 10.1186/s13662-020-02964-8
|
| [27] |
F. A. Rihan, H. J. Alsakaji, Dynamics of a stochastic delay differential model for COVID-19 infection with asymptomatic infected and interacting people: Case study in the UAE, Results Phys., 28 (2021), 104658. https://doi.org/10.1016/j.rinp.2021.104658 doi: 10.1016/j.rinp.2021.104658
|
| [28] |
D. Li, X. Cui, Dynamics of virus infection model with nonlytic immune response induced by stochastic noise, Chaos Solitons Fractals, 99 (2017), 124–132. https://doi.org/10.1016/j.chaos.2017.03.068 doi: 10.1016/j.chaos.2017.03.068
|
| [29] |
A. Din, Y. Li, T. Khan, K. Anwar, G. Zaman, Stochastic dynamics of hepatitis B epidemics, Results Phys., 20 (2021), 103730. https://doi.org/10.1016/j.rinp.2020.103730 doi: 10.1016/j.rinp.2020.103730
|
| [30] |
A. Din, A. Khan, D. Baleanu, Stationary distribution and extinction of stochastic coronavirus (COVID-19) epidemic model, Chaos Solitons Fractals, 139 (2020), 110036. https://doi.org/10.1016/j.chaos.2020.110036 doi: 10.1016/j.chaos.2020.110036
|
| [31] |
A. Din, Y. Li, Stochastic optimal control for norovirus transmission dynamics by contaminated food and water, Chin. Phys. B, 31 (2022), 020202. https://doi.org/10.1088/1674-1056/ac2f32 doi: 10.1088/1674-1056/ac2f32
|
| [32] |
C. Rajivganthi, F. A. Rihan, Global dynamics of a stochastic viral infection model with latently infected cells, Appl. Sci., 11 (2021), 10484. https://doi.org/10.3390/app112110484 doi: 10.3390/app112110484
|
| [33] |
N. Dalal, D. Greenhalgh, X. Mao, A stochastic model for internal HIV dynamics, J. Math. Anal. Appl., 341 (2008), 1084–1101. https://doi.org/10.1016/j.jmaa.2007.11.005 doi: 10.1016/j.jmaa.2007.11.005
|
| [34] |
M. Mahrouf, M. Lotfi, M. Maziane, K. Hattaf, N. Yousfi, Stability analysis for stochastic differential equations in virology, J. Adv. Math. Comput. Sci., 20 (2017), 1–12. https://doi.org/10.9734/BJMCS/2017/30345 doi: 10.9734/BJMCS/2017/30345
|
| [35] |
M. Mahrouf, K. Hattaf, N. Yousfi, Dynamics of a stochastic viral infection model with immune response, Math. Modell. Nat. Phenom., 12 (2017), 15–32. https://doi.org/10.1051/mmnp/201712502 doi: 10.1051/mmnp/201712502
|
| [36] |
F. Xie, M. Shan, X. Lian, W. Wang, Periodic solution of a stochastic HBV infection model with logistic hepatocyte growth, Appl. Math. Comput., 293 (2017), 630–641. https://doi.org/10.1016/j.amc.2016.06.028 doi: 10.1016/j.amc.2016.06.028
|
| [37] |
F. A. Rihan, H. J. Alsakaji, Analysis of a stochastic HBV infection model with delayed immune response, Math. Biosci. Eng., 18 (2021), 5194–5220. https://doi.org/10.3934/mbe.2021264 doi: 10.3934/mbe.2021264
|
| [38] |
M. E. Fatini, D. Bouggar, I. Sekkak, A. Laaribi, Stochastic near-optimal control for drug therapy in a random viral model with cellular immune response, Stochastic Anal. Appl., 4 (2021), 1–27. https://doi.org/10.1080/07362994.2021.1882312 doi: 10.1080/07362994.2021.1882312
|
| [39] |
Q. Liu, D. Jiang, N. Shi, T. HAYAT, A. ALSAEDI, Dynamical behavior of a stochastic HBV infection model with logistic hepatocyte growth, Acta Math. Sci., 37 (2017), 927–940. https://doi.org/10.1016/S0252-9602(17)30048-6 doi: 10.1016/S0252-9602(17)30048-6
|
| [40] |
H. Hui, L. Nie, Analysis of a stochastic HBV infection model with nonlinear incidence rate, J. Biol. Syst., 27 (2019), 399–421. https://doi.org/10.1142/S0218339019500177 doi: 10.1142/S0218339019500177
|
| [41] |
K. Akdim, A. Ez-zetouni, J. Danane, K. Allali, Stochastic viral infection model with lytic and nonlytic immune responses driven by Lévy noise, Physica A, 549 (2020), 124367. https://doi.org/10.1016/j.physa.2020.124367 doi: 10.1016/j.physa.2020.124367
|
| [42] |
Y. Yuan, L. J. Allen, Stochastic models for virus and immune system dynamics, Math. Biosci., 234 (2011), 84–94. https://doi.org/10.1016/j.mbs.2011.08.007 doi: 10.1016/j.mbs.2011.08.007
|
| [43] | X. Mao, Stochastic Differential Equations and Applications, Elsevier, 2007. |
| [44] |
D. J. Higham, An algorithmic introduction to numerical simulation of stochastic differential equations, SIAM Rev., 43 (2001), 525–546. https://doi.org/10.1137/s0036144500378302 doi: 10.1137/s0036144500378302
|
| [45] |
Y. Wang, Y. Zhou, Mathematical modeling and dynamics of HIV progression and treatment, Chin. J. Eng. Math., 27 (2010), 534–548. https://doi.org/10.3788/gzxb20103904.0680 doi: 10.3788/gzxb20103904.0680
|


Figures(3) / Tables(1)

Yiping Tan, Yongli Cai, Zhihang Peng, Kaifa Wang, Ruoxia Yao, Weiming Wang. Dynamics of a stochastic HBV infection model with drug therapy and immune response[J]. Mathematical Biosciences and Engineering, 2022, 19(8): 7570-7585. doi: 10.3934/mbe.2022356







 DownLoad:
DownLoad: