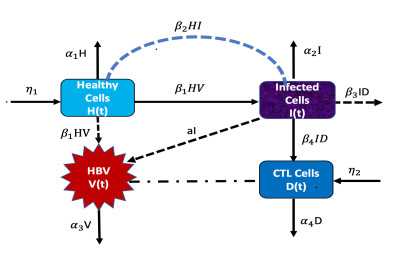
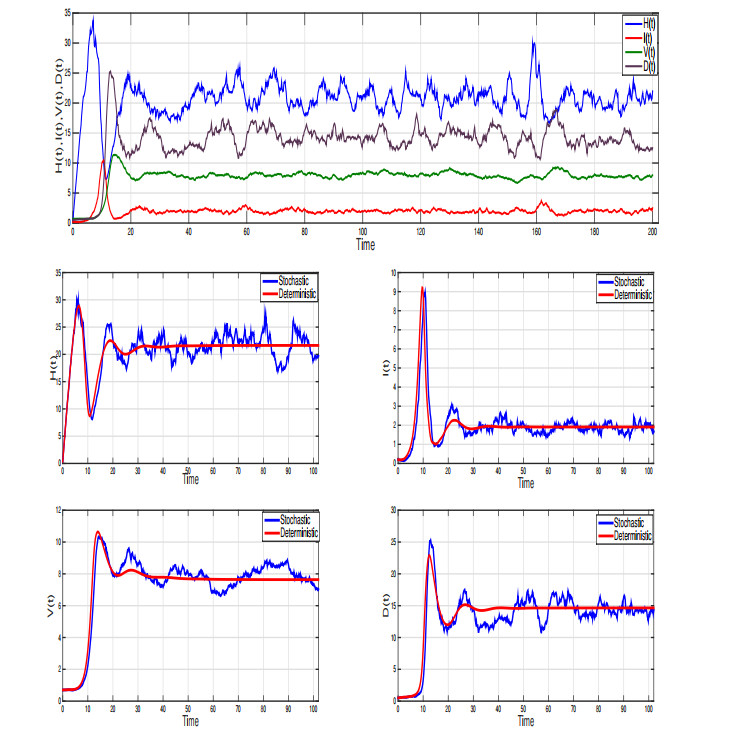
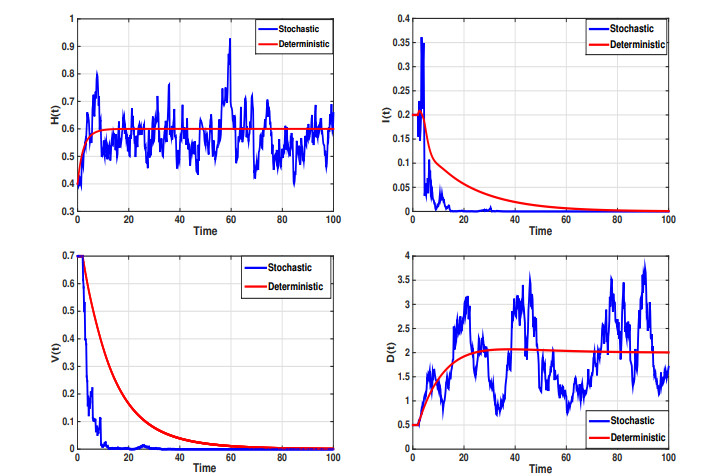
Considering the environmental factors and uncertainties, we propose, in this paper, a higher-order stochastically perturbed delay differential model for the dynamics of hepatitis B virus (HBV) infection with immune system. Existence and uniqueness of an ergodic stationary distribution of positive solution to the system are investigated, where the solution fluctuates around the endemic equilibrium of the deterministic model and leads to the stochastic persistence of the disease. Under some conditions, infection-free can be obtained in which the disease dies out exponentially with probability one. Some numerical simulations, by using Milstein's scheme, are carried out to show the effectiveness of the obtained results. The intensity of white noise plays an important role in the treatment of infectious diseases.
Citation: Fathalla A. Rihan, Hebatallah J. Alsakaji. Analysis of a stochastic HBV infection model with delayed immune response[J]. Mathematical Biosciences and Engineering, 2021, 18(5): 5194-5220. doi: 10.3934/mbe.2021264
Considering the environmental factors and uncertainties, we propose, in this paper, a higher-order stochastically perturbed delay differential model for the dynamics of hepatitis B virus (HBV) infection with immune system. Existence and uniqueness of an ergodic stationary distribution of positive solution to the system are investigated, where the solution fluctuates around the endemic equilibrium of the deterministic model and leads to the stochastic persistence of the disease. Under some conditions, infection-free can be obtained in which the disease dies out exponentially with probability one. Some numerical simulations, by using Milstein's scheme, are carried out to show the effectiveness of the obtained results. The intensity of white noise plays an important role in the treatment of infectious diseases.

| [1] |
S. M. Ciupe, R. M. Ribeiro, P. W. Nelson, A. S. Perelson, Modeling the mechanisms of acute hepatitis B virus infection, J. Theor. Biol., 247 (2007), 23-35. doi: 10.1016/j.jtbi.2007.02.017

|
| [2] |
R. M. Ribeiro, A. Lo, A. S. Perelson, Dynamics of hepatitis B virus infection, Microb. Infect., 4 (2002), 829-835. doi: 10.1016/S1286-4579(02)01603-9
|
| [3] | J. I. Weissberg, L. L. Andres, C. I. Smith, S. Weick, J. E. Nichols, G. Garcia, et al, Survival in chronic hepatitis B, Ann. Intern. Med., 101 (5), 613-616. |
| [4] |
I. S. Oh, S. H. Park, Immune-mediated liver injury in hepatitis B virus infection, Immun. Netw., 15 (2015), 191. doi: 10.4110/in.2015.15.4.191
|
| [5] | C. A. Janeway, J. P. Travers, M. Walport, M. J. Sholmchik, Immunobiology: The Immune System in Health and Disease 5th edition, New York, Garland Science, 2001. |
| [6] | F. A. Rihan, Delay Differential Equations and Applications to Biology, 2021. |
| [7] |
K. Hattaf, N. Yousfi, A generalized HBV model with diffusion and two delays, Comput. Math. Appl., 69 (2015), 31-40. doi: 10.1016/j.camwa.2014.11.010
|
| [8] |
K. Manna, S. P. Chakrabarty, Global stability of one and two discrete delay models for chronic hepatitis B infection with {HBV DNA}-containing capsids, Comput. Appl. Math., 36 (2017), 525-536. doi: 10.1007/s40314-015-0242-3
|
| [9] | K. Hattaf, K. Manna, Modeling the dynamics of hepatitis B virus infection in presence of capsids and immunity, in Mathematical Modelling and Analysis of Infectious Diseases, (2020), 269-294. |
| [10] |
T. Luzyanina, G. Bocharov, Stochastic modeling of the impact of random forcing on persistent hepatitis B virus infection, Math. Comput. Simul., 96 (2014), 54-65. doi: 10.1016/j.matcom.2011.10.002
|
| [11] |
X. Wang, Y. Tan, Y. Cai, K. Wang, W. Wang, Dynamics of a stochastic HBV infection model with cell-to-cell transmission and immune response, Math. Biosci. Eng., 18 (2021), 616-642. doi: 10.3934/mbe.2021034
|
| [12] |
C. Ji, The stationary distribution of hepatitis B virus with stochastic perturbation, Appl. Math. Lett., 100 (2020), 106017. doi: 10.1016/j.aml.2019.106017
|
| [13] |
T. Khan, I. H. Jung, G. Zaman, A stochastic model for the transmission dynamics of hepatitis B virus, J. Biol. Dyn., 13 (2019), 328-344. doi: 10.1080/17513758.2019.1600750
|
| [14] | D. Kiouach, Y. Sabbar, Ergodic stationary distribution of a stochastic hepatitis B epidemic model with interval-valued parameters and compensated poisson process, Comput. Math. Methods Med., 2020 (2020). |
| [15] | G. Bocharov, V. Volpert, B. Ludewig, A. Meyerhans, Mathematical Immunology of Virus Infections, 2018. |
| [16] |
I. Sazonov, D. Grebennikov, M. Kelbert, G. Bocharov, Modelling stochastic and deterministic behaviours in virus infection dynamics, Math. Model Nat. Phenom., 12 (2017), 63-77. doi: 10.1051/mmnp/201712505
|
| [17] | Y. Yang, L. Zou, S. Ruan, Global Dynamics of a Delayed Within-Host Viral Infection Model with Both Virus-to-Cell and Cell-to-Cell Transmissions, 2015. |
| [18] | S. Hews, S. Eikenberry, J. D. Nagy, Y. Kuang, Rich Dynamics of a Hepatitis B Viral Infection Model with Logistic Hepatocyte Growth, 2010. |
| [19] |
Y. Wang, Z. Du, W. R. Lawrence, Y. Huang, Y. Deng, Y. Hao, Predicting hepatitis B virus infection based on health examination data of community population, Int. J. Environ. Res. Public Health, 16 (2019), 4842. doi: 10.3390/ijerph16234842
|
| [20] |
X. Lai, X. Zou, Modeling HIV-1 virus dynamics with both virus-to-cell infection and cell-to-cell transmission, SIAM J. Appl. Math., 74 (2014), 898-917. doi: 10.1137/130930145
|
| [21] |
F. A. Rihan, G. Velmurugan, Dynamics and sensitivity analysis of fractional-order delay differential model for coronavirus (COVID-19) infection, Prog. Fract. Differ. Appl., 7 (2021), 43-61. doi: 10.18576/pfda/070105
|
| [22] |
S. Pan, S. P. Chakrabarty, Threshold dynamics of HCV model with cell-to-cell transmission and a non-cytolytic cure in the presence of humoral immunity, Commun. Nonlinear Sci. Numer. Simul., 61 (2018), 180-197. doi: 10.1016/j.cnsns.2018.02.010
|
| [23] |
F. A. Rihan, M. Sheek-Hussein, A. Tridane, R. Yafia, Dynamics of hepatitis C virus infection: mathematical modeling and parameter estimation, Math. Model Nat. Phenom., 12 (2017), 33-47. doi: 10.1051/mmnp/201712503
|
| [24] |
A. Goyal, L. E. Liao, A. S. Perelson, Within-host mathematical models of hepatitis B virus infection: Past, present, and future, Curr. Opin. Syst. Biol., 18 (2019), 27-35. doi: 10.1016/j.coisb.2019.10.003
|
| [25] | M. Nowak, R. M. May, Virus Dynamics: Mathematical Principles of Immunology and Virology, Oxford University Press, 2000. |
| [26] | X. Zhang, H. Peng, Stationary distribution of a stochastic cholera epidemic model with vaccination under regime switching, Appl. Math. Lett., 102 (2020). |
| [27] |
F. A. Rihan, H. J. Alsakaji, C. Rajivganthi, Stochastic SIRC epidemic model with time-delay for COVID-19, Adv. Differ. Equation, 2020 (2020), 1-20. doi: 10.1186/s13662-019-2438-0
|
| [28] |
Q. Liu, D. Jiang, T. Hayat, A. Alsaedi, B. Ahmad, Dynamical behavior of a higher order stochastically perturbed SIRI epidemic model with relapse and media coverage, Chaos Solitons Fractals., 139 (2020), 110013. doi: 10.1016/j.chaos.2020.110013
|
| [29] | X. Mao, Stochastic Differential Equations and Their Applications, Horwood, Chichester, 1997. |
| [30] |
Q. Liu, D. Jiang, Stationary distribution and extinction of a stochastic SIR model with nonlinear perturbation, Appl. Math. Lett., 73 (2017), 8-15. doi: 10.1016/j.aml.2017.04.021
|
| [31] |
Q. Liu, D. Jiang, T. Hayat, B. Ahmad, Asymptotic behavior of a stochastic delayed HIV-1 infection model with nonlinear incidence, Phys. A, 486 (2017), 867-882. doi: 10.1016/j.physa.2017.05.069
|
| [32] | R. Z. Hasminskii, Stochastic Stability of Differential Equations, Alphen aan den Rijn, Sijthoff & Noordhoff, 1980. |
| [33] |
S. Pengd, X. Zhu, Necessary and sufficient condition for comparison theorem of 1-dimensional stochastic differential equations, Stoch. Process. Their. Appl., 116 (2006), 370-380. doi: 10.1016/j.spa.2005.08.004
|
| [34] | A. Berman, R. Plemmons, Nonnegative Matrices in the Mathematical Sciences, SIAM, 1994. |
| [35] | R. S. Liptser, A strong law of large numbers for local martingales, Stochastics, 3247 (1980), 217-228. |
| [36] | X. Mao, Stochastic Differential Equations and Applications, Elsevier, 2007. |
| [37] |
H. Dahari, A. Lo, R. M. Ribeiro, A. S. Perelson, Modeling hepatitis C virus dynamics: Liver regeneration and critical drug efficacy, J. Theor. Biol., 247 (2007), 371-381. doi: 10.1016/j.jtbi.2007.03.006
|
| [38] |
D. Wodarz, Mathematical models of immune effector responses to viral infections: Virus control versus the development of pathology, J. Comput. Appl. Math., 184 (2005), 301-319. doi: 10.1016/j.cam.2004.08.016
|
| [39] |
J. Reyes-Silveyra, A. R. Mikler, Modeling immune response and its effect on infectious disease outbreak dynamics, Theor. Biol. Med. Model., 13 (2016), 1-21. doi: 10.1186/s12976-016-0027-4
|
| [40] |
D. Wodarz, Hepatitis C virus dynamics and pathology: the role of CTL and antibody responses, J. Gen. Virol., 84 (2003), 1743-1750. doi: 10.1099/vir.0.19118-0
|
| [41] |
C. Bake, E. Buckwar, Numerical analysis of explicit one-step methods for stochastic delay differential equations, LMS J. Comput. Math., 3 (2000), 315-335. doi: 10.1112/S1461157000000322
|
| [42] |
Z. Wang, C. Zhang, An analysis of stability of Milstein method for stochastic differential equations with delay, Comput. Math. Appl., 51 (2006), 1445-1452. doi: 10.1016/j.camwa.2006.01.004
|
| [43] | B. Berrhazi, M. E. Fatini, T. G. Caraballo, R. Pettersson, A stochastic SIRI epidemic model with lévy noise, Discret. Contin. Dyn. Syst. Ser. B., 23 (2018), 3645-3661. |
| [44] |
K. Hattaf, A new generalized definition of fractional derivative with non-singular kernel, Computation, 8 (2020), 49. doi: 10.3390/computation8020049
|
| [45] |
F. A. Rihan, A. A. Arafa, R. Rakkiyappand, S. Rajivganthi, Y. Xu, Fractional-order delay differential equations for the dynamics of hepatitis C virus infection with IFN-$\alpha$ treatment, Alex. Eng. J., 60 (2021), 4761-4774. doi: 10.1016/j.aej.2021.03.057
|
| [46] |
J. M. Heffernan, R. J. Smith, L. M. Wahl, Perspectives on the basic reproductive ratio, J. R. Soc. Interf., 2 (2005), 281-293. doi: 10.1098/rsif.2005.0042
|
Figures(3) / Tables(2)

Fathalla A. Rihan, Hebatallah J. Alsakaji. Analysis of a stochastic HBV infection model with delayed immune response[J]. Mathematical Biosciences and Engineering, 2021, 18(5): 5194-5220. doi: 10.3934/mbe.2021264







 DownLoad:
DownLoad: