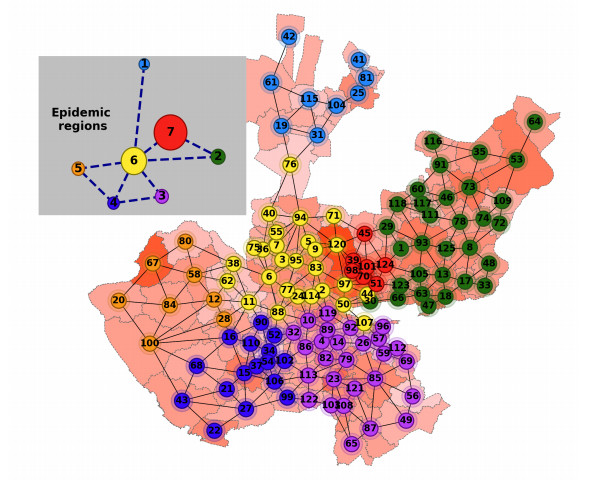
Pandemic due to SARS-CoV-2 (COVID-19) has affected to world in several aspects: high number of confirmed cases, high number of deaths, low economic growth, among others. Understanding of spatio-temporal dynamics of the virus is helpful and necessary for decision making, for instance to decide where, whether and how, non-pharmaceutical intervention policies are to be applied. This point has not been properly addressed in literature since typical strategies do not consider marked differences on the epidemic spread across country or large territory. Those strategies assume similarities and apply similar interventions instead. This work is focused on posing a methodology where spatio-temporal epidemic dynamics is captured by means of dividing a territory in time-varying epidemic regions, according to geographical closeness and infection level. In addition, a novel Lagrangian-SEIR-based model is posed for describing the dynamic within and between those regions. The capabilities of this methodology for identifying local outbreaks and reproducing the epidemic curve are discussed for the case of COVID-19 epidemic in Jalisco state (Mexico). The contagions from July 31, 2020 to March 31, 2021 are analyzed, with monthly adjustments, and the estimates obtained at the level of the epidemic regions present satisfactory results since Relative Root Mean Squared Error RRMSE is below 15% in most of regions, and at the level of the whole state outstanding with RRMSE below 5%.
Citation: Roman Zúñiga Macías, Humberto Gutiérrez-Pulido, Edgar Alejandro Guerrero Arroyo, Abel Palafox González. Geographical network model for COVID-19 spread among dynamic epidemic regions[J]. Mathematical Biosciences and Engineering, 2022, 19(4): 4237-4259. doi: 10.3934/mbe.2022196
Pandemic due to SARS-CoV-2 (COVID-19) has affected to world in several aspects: high number of confirmed cases, high number of deaths, low economic growth, among others. Understanding of spatio-temporal dynamics of the virus is helpful and necessary for decision making, for instance to decide where, whether and how, non-pharmaceutical intervention policies are to be applied. This point has not been properly addressed in literature since typical strategies do not consider marked differences on the epidemic spread across country or large territory. Those strategies assume similarities and apply similar interventions instead. This work is focused on posing a methodology where spatio-temporal epidemic dynamics is captured by means of dividing a territory in time-varying epidemic regions, according to geographical closeness and infection level. In addition, a novel Lagrangian-SEIR-based model is posed for describing the dynamic within and between those regions. The capabilities of this methodology for identifying local outbreaks and reproducing the epidemic curve are discussed for the case of COVID-19 epidemic in Jalisco state (Mexico). The contagions from July 31, 2020 to March 31, 2021 are analyzed, with monthly adjustments, and the estimates obtained at the level of the epidemic regions present satisfactory results since Relative Root Mean Squared Error RRMSE is below 15% in most of regions, and at the level of the whole state outstanding with RRMSE below 5%.

| [1] | E. Dong, H. Du, L. Gardner, An interactive web-based dashboard to track COVID-19 in real time, Lancet Infect. Dis., 20 (2020), 533–534. |
| [2] | The World Bank, World Bank open data, 2021. Available from: https://databank.worldbank.org/home.aspx. |
| [3] | SEGOB, Acuerdo por el que se establecen acciones extraordinarias para atender la emergencia sanitaria generada por el virus SARS-CoV2, 2020. Available from: https://dof.gob.mx/nota_detalle.php?codigo=5590914&fecha=31%2F03%2F2020. |
| [4] | INEGI, Censo de población y vivienda 2020, características de las localidades, tabulados predefinidos, 2021. Available from: https://www.inegi.org.mx/programas/ccpv/2020/. |
| [5] |
I. Ibarra-Nava, J. A. Cardenas-De La Garza, R. E. Ruiz-Lozano, R. G. Salazar-Montalvo, Mexico and the COVID-19 response, Disaster Med. Public Health Prep., 14 (2020), e17–e18. https://doi.org/10.1017/dmp.2020.260 doi: 10.1017/dmp.2020.260

|
| [6] |
F. Knaul, H. Arreola-Ornelas, T. Porteny, M. Touchton, M. Sánchez-Talanquer, Ó. Méndez, et al., Not far enough: Public health policies to combat COVID-19 in Mexico's states, PLoS One, 16 (2021), e0251722. https://doi.org/10.1371/journal.pone.0251722 doi: 10.1371/journal.pone.0251722
|
| [7] | M. Sánchez-Talanquer, E. González-Pier, J. Sepúlveda, L. Abascal-Miguel, J. Fieldhouse, C. Del Rio, et al., Mexico's Response to COVID-19: A Case Study, Institute for Global Health Sciences, University of California, San Francisco, 2021. Available from: https://globalhealthsciences.ucsf.edu/sites/globalhealthsciences.ucsf.edu/files/mexico-covid-19-case-study-english.pdf. |
| [8] |
I. Franch-Pardo, B. M. Napoletano, F. Rosete-Verges, L. Billa, Spatial analysis and GIS in the study of COVID-19. A review, Sci. Total Environ., 739 (2020), 140033. https://doi.org/10.1016/j.scitotenv.2020.140033 doi: 10.1016/j.scitotenv.2020.140033
|
| [9] |
C. Scarpone, S. T. Brinkmann, T. Große, D. Sonnenwald, M.Fuchs, B. B. Walker, A multimethod approach for county-scale geospatial analysis of emerging infectious diseases: a cross-sectional case study of COVID-19 incidence in Germany, Int. J. Health Geographics, 19 (2020), 1–17. https://doi.org/10.1186/s12942-020-00225-1 doi: 10.1186/s12942-020-00225-1
|
| [10] |
N. Arinaminpathy, J. Das, T. H. McCormick, P. Mukhopadhyay, N. Sircar, Quantifying heterogeneity in SARS-CoV-2 transmission during the lockdown in India, Epidemics, 36 (2021), 100477. https://doi.org/10.1101/2020.09.10.20190017 doi: 10.1101/2020.09.10.20190017
|
| [11] |
Y. Ge, Z. Chen, A. Handel, L. Martinez, Q. Xiao, C. Li, et al., The impact of social distancing, contact tracing, and case isolation interventions to suppress the COVID-19 epidemic: A modeling study, Epidemics, 36 (2021), 100483. https://doi.org/10.1016/j.epidem.2021.100483 doi: 10.1016/j.epidem.2021.100483
|
| [12] |
S. Triambak, D. P. Mahapatra, N. Mallick, R. Sahoo, A new logistic growth model applied to COVID-19 fatality data, Epidemics, (2021), 100515. https://doi.org/10.1016/j.epidem.2021.100515 doi: 10.1016/j.epidem.2021.100515
|
| [13] |
P. Pincheira-Brown, A. Bentancor, Forecasting COVID-19 infections with the semi-unrestricted generalized growth model, Epidemics, 37 (2021), 100486. https://doi.org/10.1016/j.epidem.2021.100486 doi: 10.1016/j.epidem.2021.100486
|
| [14] |
M. Fatima, K. J. O'Keefe, W. Wei, S. Arshad, O. Gruebner, Geospatial analysis of COVID-19: A scoping review, Int. J. Environ. Res. Public Health, 18 (2021), 2336. https://doi.org/10.3390/ijerph18052336 doi: 10.3390/ijerph18052336
|
| [15] |
S. He, S. Tang, L. Rong, A discrete stochastic model of the COVID-19 outbreak: Forecast and control, Math. Biosci. Eng., 17 (2020), 2792–2804. https://doi.org/10.3934/mbe.2020153 doi: 10.3934/mbe.2020153
|
| [16] |
A. K. Srivastav, P. K. Tiwari, P. K. Srivastava, M. Ghosh, Y. Kang, A mathematical model for the impacts of face mask, hospitalization and quarantine on the dynamics of COVID-19 in India: deterministic vs. stochastic, Math. Biosci. Eng., 18 (2021), 182–213. https://doi.org/10.3934/mbe.2021010 doi: 10.3934/mbe.2021010
|
| [17] |
D. Olabode, J. Culp, A. Fisher, A. Tower, D. Hull-Nye, X. Wang, Deterministic and stochastic models for the epidemic dynamics of COVID-19 in Wuhan, China, Math. Biosci. Eng., 18 (2021), 950–967. https://doi.org/10.3934/mbe.2021050 doi: 10.3934/mbe.2021050
|
| [18] |
T. S. Brown, K. Engø-Monsen, M. V. Kiang, A. S. Mahmud, R. J. Maude, C. O. Buckee, The impact of mobility network properties on predicted epidemic dynamics in Dhaka and Bangkok, Epidemics, 35 (2021), 100441. https://doi.org/10.1016/j.epidem.2021.100441 doi: 10.1016/j.epidem.2021.100441
|
| [19] |
W. Chiang, X. Liu, G. Mohler, Hawkes process modeling of COVID-19 with mobility leading indicators and spatial covariates, Int. J. Forecast., 38 (2021), 505–520. https://doi.org/10.1016/j.ijforecast.2021.07.001 doi: 10.1016/j.ijforecast.2021.07.001
|
| [20] |
R. Engbert, M. M. Rabe, R. Kliegl, S. Reich, Sequential data assimilation of the stochastic SEIR epidemic model for regional COVID-19 dynamics, Bull. Math. Biol., 83 (2021), 1–16. https://doi.org/10.1007/s11538-020-00834-8 doi: 10.1007/s11538-020-00834-8
|
| [21] |
V. Gopalakrishnan, S. Pethe, S. Kefayati, S. Raman, P. Hake, A. Deshpande, et al., Globally local: Hyper-local modeling for accurate forecast of COVID-19, Epidemics, 37 (2021), 100510. https://doi.org/10.1016/j.epidem.2021.100510 doi: 10.1016/j.epidem.2021.100510
|
| [22] |
R. Huang, M. Liu, Y. Ding, Spatial-temporal distribution of COVID-19 in China and its prediction: A data-driven modeling analysis, J. Infect. Dev. Countries, 14 (2020), 246–253. https://doi.org/10.3855/jidc.12585 doi: 10.3855/jidc.12585
|
| [23] |
Y. Mammeri, A reaction-diffusion system to better comprehend the unlockdown: Application of SEIR-type model with diffusion to the spatial spread of COVID-19 in France, Comput. Math. Biophys., 8 (2020), 102–113. https://doi.org/10.1515/cmb-2020-0104 doi: 10.1515/cmb-2020-0104
|
| [24] |
X. Hou, S. Gao, Q. Li, Y. Kang, N. Chen, K. Chen, et al., Intracounty modeling of COVID-19 infection with human mobility: Assessing spatial heterogeneity with business traffic, age, and race, Proc. Natl. Acad. Sci., 118 (2021). https://doi.org/10.1073/pnas.2020524118 doi: 10.1073/pnas.2020524118
|
| [25] |
T. W. Alleman, J. Vergeynst, L. De Visscher, M. Rollier, E. Torfs, I. Nopens, et al., Assessing the effects of non-pharmaceutical interventions on SARS-CoV-2 transmission in Belgium by means of an extended SEIQRD model and public mobility data, Epidemics, 37 (2021), 100505. https://doi.org/10.1016/j.epidem.2021.100505 doi: 10.1016/j.epidem.2021.100505
|
| [26] |
R. Pastor-Satorras, C. Castellano, P. Van Mieghem, A. Vespignani, Epidemic processes in complex networks, Rev. Mod. Phys., 87 (2015), 925. https://doi.org/10.1103/RevModPhys.87.925 doi: 10.1103/RevModPhys.87.925
|
| [27] |
Y. Wang, J. Cao, Final size of network epidemic models: properties and connections, Sci. China Inf. Sci, 64 (2021), 179201. https://doi.org/10.1007/s11432-019-2656-2 doi: 10.1007/s11432-019-2656-2
|
| [28] |
D. O'Sullivan, M. Gahegan, D. J. Exeter, B. Adams, Spatially explicit models for exploring COVID-19 lockdown strategies, Trans. GIS, 24 (2020), 967–1000. https://doi.org/10.1111/tgis.12660 doi: 10.1111/tgis.12660
|
| [29] |
D. Laroze, E. Neumayer, T. Plümper, COVID-19 does not stop at open borders: Spatial contagion among local authority districts during England's first wave, Social Sci. Med., 270 (2021). https://doi.org/10.1016/j.socscimed.2020.113655 doi: 10.1016/j.socscimed.2020.113655
|
| [30] |
I. Franch-Pardo, M. R. Desjardins, I. Barea-Navarro, A. Cerdà, A review of GIS methodologies to analyze the dynamics of COVID-19 in the second half of 2020, Trans. GIS, 25 (2021), 2191–2239. https://doi.org/10.1111/tgis.12792 doi: 10.1111/tgis.12792
|
| [31] | A. Gandolfi, Percolation methods for SEIR epidemics on graphs, in Dynamic Models of Infectious Diseases, Springer, New York, NY, (2013), 31–58. https://doi.org/10.1007/978-1-4614-9224-5_2 |
| [32] | R. Wallace, L. Bergmann, L. Hogerwerf, R. Kock, R. G. Wallace, Ebola in the hog sector: Modeling pandemic emergence in commodity livestock, in Neoliberal Ebola, Springer, Cham, (2016), 13–53. https://doi.org/10.1007/978-3-319-40940-5_2 |
| [33] |
R. Wallace, D. Wallace, H. Andrews, R. Fullilove, M. T. Fullilove, The spatiotemporal dynamics of AIDS and TB in the New York metropolitan region from a sociogeographic perspective: Understanding the linkages of central city and suburbs, Environ. Plann. A, 27 (1995), 1085–1108. https://doi.org/10.1068/a271085 doi: 10.1068/a271085
|
| [34] |
U. N. Raghavan, R. Albert, S. Kumara, Near linear time algorithm to detect community structures in large-scale networks, Phys. Rev. E, 76 (2007), 036106. https://doi.org/10.1103/PhysRevE.76.036106 doi: 10.1103/PhysRevE.76.036106
|
| [35] | M. Martcheva, An introduction to Mathematical Epidemiology, Springer, 61 (2015). https://doi.org/10.1007/978-1-4899-7612-3 |
| [36] | Center for Disease Control and Prevention, Scientific brief: SARS-CoV-2 transmission, 2022. Available from: https://www.cdc.gov/coronavirus/2019-ncov/science/science-briefs/sars-cov-2-transmission.html. |
| [37] |
J. M. Brauner, S. Mindermann, M. Sharma, D. Johnston, J. Salvatier, T. Gavenčiak, et al., Inferring the effectiveness of government interventions against COVID-19, Science, 371 (2021). https://doi.org/10.1126/science.abd9338 doi: 10.1126/science.abd9338
|
| [38] |
N. Imai, K. A. M. Gaythorpe, S. Abbott, S. Bhatia, S. van Elsland, K. Prem, et al., Adoption and impact of non-pharmaceutical interventions for COVID-19, Wellcome Open Res., 5 (2020). https://doi.org/10.12688/wellcomeopenres.15808.1 doi: 10.12688/wellcomeopenres.15808.1
|
| [39] |
N. L. Komarova, A. Azizi, D. Wodarz, Network models and the interpretation of prolonged infection plateaus in the COVID19 pandemic, Epidemics, 35 (2021), 100463. https://doi.org/10.1016/j.epidem.2021.100463 doi: 10.1016/j.epidem.2021.100463
|
| [40] |
D. Pradhan, P. Biswasroy, P. Kumar Naik, G. Ghosh, G. Rath, A review of current interventions for COVID-19 prevention, Arch. Med. Res., 51 (2020), 363–374. https://doi.org/10.1016/j.arcmed.2020.04.020 doi: 10.1016/j.arcmed.2020.04.020
|
| [41] |
K. Soltesz, F. Gustafsson, T. Timpka, J. Jaldén, C. Jidling, A. Heimerson, et al., The effect of interventions on COVID-19, Nature, 588 (2020), E26–E28. https://doi.org/10.1038/s41586-020-3025-y doi: 10.1038/s41586-020-3025-y
|
| [42] |
P. Bacchetti, N. P. Jewell, Nonparametric estimation of the incubation period of AIDS based on a prevalent cohort with unknown infection times, Biometrics, 47 (1991), 947–960. https://doi.org/10.2307/2532651 doi: 10.2307/2532651
|
| [43] |
B. Cazelles, S. Hales, Infectious diseases, climate influences, and nonstationarity, PLoS Med., 3 (2006), e328. https://doi.org/10.1371/journal.pmed.0030328 doi: 10.1371/journal.pmed.0030328
|
| [44] |
J. Lu, A. Lin, C. Jiang, A. Zhang, Z. Yang, Influence of transportation network on transmission heterogeneity of COVID-19 in China, Transp. Res. Part C Emerging Technol., 129 (2021), 103231. https://doi.org/10.1016/j.trc.2021.103231 doi: 10.1016/j.trc.2021.103231
|
| [45] | F. Brauer, P. V. den Driessche, J. Wu, Mathematical Epidemiology, Berlin, Germany, Springer, 1945 (2008). https://doi.org/10.1007/978-3-540-78911-6 |
| [46] | J. Chen, S. Levin, S. Eubank, H. Mortveit, S. Venkatramanan, A. Vullikanti, et al., Networked epidemiology for COVID-19, SIAM News, 53 (2020). Available from: https://par.nsf.gov/servlets/purl/10213734. |
| [47] |
M. Marathe, A. K. S. Vullikanti, Computational epidemiology, Commun. ACM, 56 (2013), 88–96. https://doi.org/10.1145/2483852.2483871 doi: 10.1145/2483852.2483871
|
| [48] |
W. O. Kermack, A. G. McKendrick, A contribution to the mathematical theory of epidemics, Proc. R. Soc. London, Ser. A, 115 (1927), 700–721. https://doi.org/10.1098/rspa.1927.0118 doi: 10.1098/rspa.1927.0118
|
| [49] |
F. Arrigoni, A. Pugliese, Limits of a multi-patch SIS epidemic model, J. Math. Biol., 45 (2002), 419–440. https://doi.org/10.1007/s002850200155 doi: 10.1007/s002850200155
|
| [50] |
C. Wolf, M. Langlais, D. Ponter, A multi-patch epidemic model with periodic demography, direct and indirect transmission and variable maturation rate, Math. Popul. Stud., 13 (2006), 153–177. https://doi.org/10.1080/08898480600788584 doi: 10.1080/08898480600788584
|
| [51] |
J. Arino, J. M. Miller, P. Van den Driessche, A multi-species epidemic model with spatial dynamics, Math. Med. Biol., 22 (2005), 129–142. https://doi.org/10.1093/imammb/dqi003 doi: 10.1093/imammb/dqi003
|
| [52] |
R. K. McCormack, L. J. S. Allen, Multi-patch deterministic and stochastic models for wildlife diseases, J. Biol. Dyn., 1 (2007), 63–85. https://doi.org/10.1080/17513750601032711 doi: 10.1080/17513750601032711
|
| [53] |
D. Bichara, A. Iggidr, Multi-patch and multi-group epidemic models: a new framework, J. Math. Biol, 77 (2018), 107–134. https://doi.org/10.1007/s00285-017-1191-9 doi: 10.1007/s00285-017-1191-9
|
| [54] |
M. Girvan, M. E. J. Newman, Community structure in social and biological networks, Proc. Natl. Acad. Sci., 99 (2002), 7821–7826. https://doi.org/10.1073/pnas.122653799 doi: 10.1073/pnas.122653799
|
| [55] | F. Parés, D. G. Gasulla, A. Vilalta, J. Moreno, E. Ayguadé, J. Labarta, et al., Fluid communities: A competitive, scalable and diverse community detection algorithm, in International Conference on Complex Networks and Their Applications, Springer, Cham, (2017), 229–240. https://doi.org/10.1007/978-3-319-72150-7_19 |
| [56] |
S. Fortunato, Community detection in graphs, Phys. Rep., 486 (2010), 75–174. https://doi.org/10.1016/j.physrep.2009.11.002 doi: 10.1016/j.physrep.2009.11.002
|
| [57] |
A. Clauset, M. E. J. Newman, C. Moore, Finding community structure in very large networks, Phys. Rev. E, 70 (2004), 066111. https://doi.org/10.1103/PhysRevE.70.066111 doi: 10.1103/PhysRevE.70.066111
|
| [58] | M. Bastian, S. Heymann, M. Jacomy, Gephi: An open source software for exploring and manipulating networks, in Proceedings of the Third International Conference on Weblogs and Social Media, 2009. https://doi.org/10.13140/2.1.1341.1520 |
| [59] |
R. Wallace, Y. S. Huang, P. Gould, D. Wallace, The hierarchical diffusion of AIDS and violent crime among US metropolitan regions: inner-city decay, stochastic resonance and reversal of the mortality transition, Social Sci. Med., 44 (1997), 935–947. https://doi.org/10.1016/s0277-9536(96)00197-9 doi: 10.1016/s0277-9536(96)00197-9
|
| [60] | Secretaría de Salud Jalisco, Datos de Radar Jalisco, 2021. Available from: https://coronavirus.jalisco.gob.mx/datos-abiertos. |
| [61] | H. Gutiérrez-Pulido, D. Gómez-Alvarez, Estudio técnico para la adecuación de las regiones de Jalisco, Technical report, Gobierno de Jalisco, 2016. Available from: https://seplan.app.jalisco.gob.mx/biblioteca/archivo/verDocumento/939. |
| [62] |
K. Ejima, K. S. Kim, C. Ludema, A. I. Bento, S. Iwanami, Y. Fujita, et al., Estimation of the incubation period of COVID-19 using viral load data, Epidemics, 35 (2021), 100454. https://doi.org/10.1016/j.epidem.2021.100454 doi: 10.1016/j.epidem.2021.100454
|
| [63] |
H. Najafimehr, K. M. Ali, S. Safari, M. Yousefifard, M. Hosseini, Estimation of basic reproduction number for COVID-19 and the reasons for its differences, Int. J. Clin. Pract., 74 (2020), e13518. https://doi.org/10.1111/ijcp.13518 doi: 10.1111/ijcp.13518
|
| [64] |
W. He, G. Y. Yi, Y. Zhu, Estimation of the basic reproduction number, average incubation time, asymptomatic infection rate, and case fatality rate for COVID-19: Meta-analysis and sensitivity analysis, J. Med. Virol., 92 (2020), 2543–2550. https://doi.org/10.1002/jmv.26041 doi: 10.1002/jmv.26041
|
| [65] |
S. Zhao, B. Tang, S. S. Musa, S. Ma, J. Zhang, M. Zeng, et al., Estimating the generation interval and inferring the latent period of COVID-19 from the contact tracing data, Epidemics, 36 (2021), 100482. https://doi.org/10.1016/j.epidem.2021.100482 doi: 10.1016/j.epidem.2021.100482
|
| [66] |
J. A. Backer, D. Klinkenberg, J. Wallinga, Incubation period of 2019 novel coronavirus (2019-NCOV) infections among travellers from Wuhan, China, 20–28 January 2020, Eurosurveillance, 25 (2020), 2000062. https://doi.org/10.2807/1560-7917.ES.2020.25.5.2000062 doi: 10.2807/1560-7917.ES.2020.25.5.2000062
|
| [67] |
R. J. Hyndman, A. B. Koehler, Another look at measures of forecast accuracy, Int. J. Forecast., 22 (2006), 679–688. https://doi.org/10.1016/j.ijforecast.2006.03.001 doi: 10.1016/j.ijforecast.2006.03.001
|
 mbe-19-04-196-supplementary.pdf mbe-19-04-196-supplementary.pdf |
 |



Figures(4)

Roman Zúñiga Macías, Humberto Gutiérrez-Pulido, Edgar Alejandro Guerrero Arroyo, Abel Palafox González. Geographical network model for COVID-19 spread among dynamic epidemic regions[J]. Mathematical Biosciences and Engineering, 2022, 19(4): 4237-4259. doi: 10.3934/mbe.2022196







 DownLoad:
DownLoad: