Citation: David Moreno Martos, Benjamin J. Parcell, Raluca Eftimie. Modelling the transmission of infectious diseases inside hospital bays: implications for COVID-19[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 8084-8104. doi: 10.3934/mbe.2020410

| [1] |
B. Carter, J. T. Collins, F. Barlow-Pay, F. Rickard, E. Bruce, et al., Nosocomial COVID-19 infection: examining the risk of mortality. The COPE-Nosocomial study (COVID in Older PEople), J. Hosp. Infect., 106 (2020), 376-384. doi: 10.1016/j.jhin.2020.07.013

|
| [2] | H. M. Rickman, T. Rampling, K. Shaw, G. Martinez-Garcia, L. Hail, P. Coen, et al., Nosocomial transmission of COVID-19: a retrospective study of 66 hospital-acquired cases in a London teaching hospital, Clin. Infect. Dis., (2020), ciaa816. |
| [3] |
Q. Zhou, Y. Gao, X. Wang, R. Liu, P. Du, X. Wang, et al., Nosocomial infections among patients with COVID-19, SARS and MERS: a rapid review and meta-analysis, Ann. Transl. Med., 8 (2020), 629. doi: 10.21037/atm-20-3324
|
| [4] | K. Mizumoto, K. Kagaya, A. Zarebski, G. Chowell, Estimating the asymptomatic proportion of coronavirus disease 2019 (COVID-19) cases on board the Diamond Princess cruise ship, Yokohama, Japan, 2020, Euro. Surveil., 25 (2020), pii = 2000180. https://doi.org/10.2807/1560-7917.ES.2020.25.10.2000180. |
| [5] |
H. Nishiura, T. Miyama, A. Suzuki, S.M. Jung, K. Hayashi, R. Kinoshita, et al., Estimation of the asymptomatic ratio of novel coronavirus infections (COVID-19), Int. J. Infect. Dis., 94 (2020), 154-155. doi: 10.1016/j.ijid.2020.03.020
|
| [6] | M. Day, COVID-19: identifying and isolating asymptomatic people helped eliminate virus in Italian village, BMJ, 368 (2020), m1165. |
| [7] |
A. J. Ing, C. Cocks, J. P. Green, COVID-19: in the footsteps of Ernest Shackleton, Thorax, 75 (2020), 693-694. doi: 10.1136/thoraxjnl-2020-215091
|
| [8] |
S. Tian, N. Hu, J. Lou, K. Chen, X. Kang, Z. Xiang, et al., Characteristics of COVID-19 infection in Beijing, J. Infect., 80 (2020), 401-406. doi: 10.1016/j.jinf.2020.02.018
|
| [9] | J. Maben, P. Griffiths, C. Penfold, M. Simon, E. Pizzo, J. Anderson, et al., Evaluating a major innovation in hospital design: workforce implications and impact on patient and staff experiences of all single room hospital accommodation, Health. Serv. Deliv. Res., 3 (2015), 1-342. |
| [10] |
X. Wang, Q. Zhou, Y. He, L. Liu, X. Ma, X. Wei, et al., Nosocomial outbreak of COVID-2019 pneumonia in Wuhan, China, Eur. Respir. J., 55 (2020), 2000544. doi: 10.1183/13993003.00544-2020
|
| [11] |
Y. Fang, Y. Nie, M. Penny, Transmission dynamics of the COVID-19 outbreak and effectiveness of government interventions: A data-driven analysis, J. Med. Virol., 92 (2020), 645-659. doi: 10.1002/jmv.25750
|
| [12] | A. Grant, Dynamics of COVID-19 epidemics: SEIR models underestimate peak infection rates and overestimate epidemic duration, medRxiv, (2020), https://doi.org/10.1101/2020.04.02.20050674. |
| [13] | A. J. Kucharski, T. W. Russell, C. Diamond, Y. Liu, J. Edmunds, S. Funk, et al., Early dynamics of transmission and control of COVID-19: a mathematical modelling study, Lancet Infect. Dis., 20 (2020), P553-558. |
| [14] | L. R. Lopez, X. Rodo, A modified SEIR model to predict the COVID-19 outbreak in Spain and Italy: simulating control scenarios and multi-scale epidemics, available at SSRN 3576802, (2020), http://dx.doi.org/10.2139/ssrn.3576802. |
| [15] | J. M. Read, J. R. E. Bridgen, D. A. T. Cummings, A. Jo, C. P. Jewell, Novel coronavirus 2019-nCoV: early estimation of epidemiological parameters and epidemic predictions, medRxiv, (2020), https://doi.org/10.1101/2020.01.23.20018549. |
| [16] | J. Arino, A simple model for COVID-19, Infect. Disease Model., 5 (2020), 309-315. |
| [17] |
D. He, S. Zhao, Q. Lin, Z. Zhuang, P. Gao, M. H. Wang, et al., The relative transmissibility of asymptomatic COVID-19 infections among close contacts, Int. J. Infect. Dis., 94 (2020), 145-147. doi: 10.1016/j.ijid.2020.04.034
|
| [18] | J. Pan, Y. Zhao, Z. Liu, M. Li, Y. Wang, W. Dong, et al., Effectiveness of control strategies for Coronavirus Disease 2019: a SEIR dynamic modelling study, Bull World Health Organ, E-pub, (2020), http://dx.doi.org/10.2471/BLT.20.253807. |
| [19] | D. W. Berger, K. F. Herkenhoff, S. Mongey, An SEIR infectious disease model with testing and conditional quarantine, Natl. Bur. Econ. Res., (2020), https://doi.org/10.3386/w26901. |
| [20] | L. Peng, W. Yang, D. Zhang, C. Zhuge, L. Hong, Epidemic analysis of COVID-19 in China by dynamical modelling, medRxiv, (2020), https://doi.org/10.1101/2020.02.16.20023465. |
| [21] | Y. Liu, Z. Gu, S. Xia, B. Shi, X.-N. Zhou, Y. Shi, et al., What are the underlying transmission patterns of COVID-19 outbreak? An age-specific social contact characterisation, Lancet EClin. Med., 22 (2020), 100354. |
| [22] | K. Prem, Y. Liu, T. W. Russell, A. J. Kucharski, R. M. Eggo, N. Davies, et al., The effect of control strategies to reduce social mixing on outcomes of the COVID-19 epidemic in Wuhan, China: a modelling study, Lancet Public Health, 5 (2020), E261-E270. |
| [23] | A. Teslya, T. M. Pham, N. G. Godijk, M. E. Kretzschmar, M. C. J. Bootsma, G. Rozhnova, Impact of self-imposed prevention measures and short-term government-imposed social distancing on mitigating and delaying a COVID-19 epidemic: A modelling study, PLoS Med., 17, (2020), e1003166. |
| [24] |
Z. Yang, Z. Xheng, K. Wang, S-S. Wong, W. Liang, M. Zanin, et al., Modified SEIR and AI prediction of the epidemics trend of COVID-19 in China under public health interventions, J. Thorac. Dis., 12 (2020), 165-174. doi: 10.21037/jtd.2020.02.64
|
| [25] | A. J. Kucharski, P. Klepac, A. J. K. Conlan, S. M. Kissler, M. L. Tang, H. Fry, et al., Effectiveness of isolation, testing, contact tracing, and physical distancing on reducing transmission of SARSCoV-2 in different settings: a mathematical modelling study, Lancet Infect. Disease., 20 (2020), P1151-1160. |
| [26] | S. Evans, E. Agnew, E. Vynnycky, J. V. Robotham, The impact of testing and infection prevention and control strategies on within-hospital transmission dynamics of COVID-19 in English hospitals, medRxiv, (2020), https://doi.org/10.1101/2020.05.12.20095562. |
| [27] | E. G. Nepomuceno, R. H. C. Takahashi, L. A. Aguirre, Individual-based model (IBM): an alternative framework for epidemiological compartment models, Rev. Bras. Biom., 34 (2016), 133-162. |
| [28] | S. Romano-Bertrand, L.-S. Aho-Glele, B. Grandbastien, J.-F. Gehanno, D. Lepelletier, Sustainability of SARS-CoV-2 in aerosols: should we worry about airborne transmission? J. Hosp. Infect., 105 (2020), 601-603. |
| [29] |
M. Klompas, M. A. Baker, C. Rhee, Airborne transmission of SARS-CoV-2. Theoretical considerations and available evidence, JAMA, 324 (2020), 441-442. doi: 10.1001/jama.2020.12458
|
| [30] | Public Health England, Reducing the risk of transmission of COVID-19 in the hospital setting, 2020. Available from: https://www.gov.uk/government/publications/wuhan-novel-coronavirus-infection-prevention-and-control/reducing-the-risk-of-transmission-of-covid-19-in-the-hospital-setting. |
| [31] | D. Moreno-Martos, Modelling COVID-19 transmission in hospital bays. MSc Thesis. University of Dundee, UK (2020). |
| [32] |
T. Chen, J. Rui, Q.-P. Wang, Z.-Y. Zhao, J.-A. Cui, L. Yin, A mathematical model for simulating the phase-based transmissibility of a novel coronavirus, Infect. Dis. Poverty, 9 (2020), 24. doi: 10.1186/s40249-020-00640-3
|
| [33] | F. B. Hamzah, C. Lau, H. Nazri, D. V. Ligot, G. Lee, C. L. Tan, et al., CoronaTracker: World-wide COVID-19 outbreak data analysis and prediction, Bull World Health Organ, 1 (2020), 32. |
| [34] | COVID-19 Coronavirus Pandemic, (2020). Available from: https://www.worldometers.info/coronavirus. |
| [35] | L. Danon, E. Brooks-Pollock, M. Bailey, M. J. Keeling, A spatial model of COVID-19 transmission in England and Wales: early spread and peak timing, medRxiv, (2020), https://doi.org/10.1101/2020.02.12.20022566. |
| [36] |
B. Tang, X. Wang, Q. Li, N. L. Bragazzi, S. Tang, Y. Xiao, et al., Estimation of the transmission risk of the 2019-nCoV and its implication for public health interventions, J. Clin. Med., 9 (2020), 462. doi: 10.3390/jcm9020462
|
| [37] | H. Wang, Z. Wang, Y. Dong, R. Chang, C. Xu, X. Yu, et al., Phase-adjusted estimation of the number of Coronavirus Disease 2019 cases in Wuhan, China, Cell Discov., 6 (2020), 10. |
| [38] |
S. A. Lauer, K. H. Grantz, Q. Bi, F. K. Jones, Q. Zheng, H. R. Meredith, et al., The Incubation Period of Coronavirus Disease 2019 (COVID-19) From Publicly Reported Confirmed Cases: Estimation and Application, Ann. Intern. Med., 172 (2020), 577-582. doi: 10.7326/M20-0504
|
| [39] |
F. Ndaïrou, I. Area, J. J. Nieto, D. F. M. Torres, Mathematical modelling of COVID-19 transmission dynamics with a case study of Wuhan, Chaos Solitons Fractals, 135 (2020), 109846. doi: 10.1016/j.chaos.2020.109846
|
| [40] | R. F. Reis, B. M. Quintela, J. O. Campos, J. M. Gomes, B. M. Rocha, M. Lobosco, et al., Characterisation of the COVID-19 pandemic and the impact of uncertainties, mitigation strategies, and underreporting of cases in South Korea, Italy, and Brazil, Chaos Solitons Fractals, 136, (2020), 109888. |
| [41] | C. Savvides, R. Siegel, Asymptomatic and presymptomatic transmission of SARS-CoV-2: A systematic review, medRxiv, (2020), doi: 10.1101/2020.06.11.20129072. |
| [42] | Z.-D. Tong, A. Tang, K.-F. Li, P. Li, H.-L. Wang, J.-P. Yi, et al., Potential presymptomatic transmission of SARS-CoV-2, Zhejiang Province, China, 2020, Emerg. Infect. Dis., 26(5) (2020), 1052-1054. |
| [43] | P. Yu, J. Zhu, Z. Zhang, Y. Han, A familial cluster of infection associated with the 2019 novel coronavirus indicating possible person-to-person transmission during the incubation period, J. Infect. Dis., 221, (2020), 1757-1761. |
| [44] |
R. S. Sikkema, S. D. Pas, D. F. Nieuwenhuijse, A. O'Toole, J. J. Verweij, A. van der Linden, et al., COVID-19 in health-care workers in three hospitals in the south of the Netherlands: a crosssectional study, Lancet Infect. Dis., 20 (2020), 1273-1280. doi: 10.1016/S1473-3099(20)30527-2
|
| [45] |
N. van Doremalen, T. Bushmaker, D. H. Morris, M. G. Holbrook, A. Gamble, B. N. Williamson, et al., Aerosol and surface stability of SARS-CoV-2 as compared with SARS-Cov-1, N. Engl. J. Med., 382 (2020), 1564-1567. doi: 10.1056/NEJMc2004973
|
| [46] | M. E. Halloran, Secondary attack rate, In Encyclopedia of Biostatistics (eds. P. Armitage and T. Colton), American Cancer Society, (2005). |
| [47] |
R. M. Burke, C. M. Midgley, A. Dratch, M. Fenstersheib, T. Haupt, M. Holshue, et al., Active monitoring of persons exposed to patients with confirmed COVID-19 - United States, JanuaryFebruary 2020, MMWR Morb. Mortal. Wkly Rep., 69 (2020), 245-246. doi: 10.15585/mmwr.mm6909e1
|
| [48] | Y. Liu, R. M. Eggo, A. J. Kucharski, Secondary attack rate and superspreading events for SARSCoV-2, Lancet, 395 (2020), e47. |
| [49] | W. W. Sun, F. Ling, J. R. Pan, J. Cai, Z. P. Miao, S. L. Liu, et al., Epidemiological characteristics of COVID-19 family clustering in Zhejiang Province, Zhonghua Yu Fang Yi Xue Za Zhi, 54 (2020), 625-629. |
| [50] | Y.-T. Huang, Y.-K. Tu, P.-C. Lai, Estimation of the secondary attack rate of COVID-19 using proportional meta-analysis of nationwide contact tracing data in Taiwan, J. Microbiol. Immun. Infect., (2020), S1684-1182(20)30143-2. |
| [51] | D. K. Chu, E. A. Akl, S. Duda, K. Solo, S. Yaacoub, H. J. Schünemann, et al., Physical distancing, face masks, and eye protection to prevent person-to-person transmission of SARS-CoV-2 and COVID-19: a systematic review and meta-analysis, Lancet, 395 (2020), P1973-1987. |
| [52] | C. R. McIntyre, Q. Wang, Physical distancing, face masks, and eye protection for prevention of COVID-19, Lancet, 395 (2020), P1950-1951. |
| [53] |
Q. X. Long, X.-J. Tang, Q.-L. Shi, Q. Lin, H.-J. Deng, J. Yuan, et al., Clinical and immunological assessment of asymptomatic SARS-CoV-2 infections, Nat. Med., 26 (2020), 1200-1204. doi: 10.1038/s41591-020-0965-6
|
| [54] | M. Day, COVID-19: four fifths of cases are asymptomatic, China figures indicate, BMJ, 369 (2020), m1375. |
| [55] |
G. Giordano, F. Blanchini, R. Bruno, P. Colaneri, A. Di Filippo, A. Di Matteo, et al., Modelling the COVID-19 epidemic and implementation of population-wide interventions in Italy, Nat. Med., 26 (2020), 855-860. doi: 10.1038/s41591-020-0883-7
|
| [56] | Health Protection Scotland, Novel coronavirus (COVID-19) guidance for secondary care, (2020). Available from: https://www.hps.scot.nhs.uk/web-resources-container/covid-19-guidance-for-secondary-care/. |
| [57] | Interim Guidance for Use of Pooling Procedures in SARS-CoV-2 Diagnostic, Screening, and Surveillance Testing, (2020). Available from: https://www.cdc.gov/coronavirus/2019-ncov/lab/pooling-procedures.html. |
| [58] | T. Roques, R. Board, Guidance on SARS-CoV-2 antigen testing for asymptomatic healthcare workers (HCW) and patients in non-surgical oncology in the UK, (2020). Available from https://www.rcr.ac.uk/sites/default/files/guidance-covid19-testing-asymptomatic-hcw-patients-oncology.pdf. |
| [59] |
C. Aitken, D. J. Jeffries, Nosocomial spread of viral disease, Clin. Microbiol. Rev., 14 (2001), 528-546. doi: 10.1128/CMR.14.3.528-546.2001
|
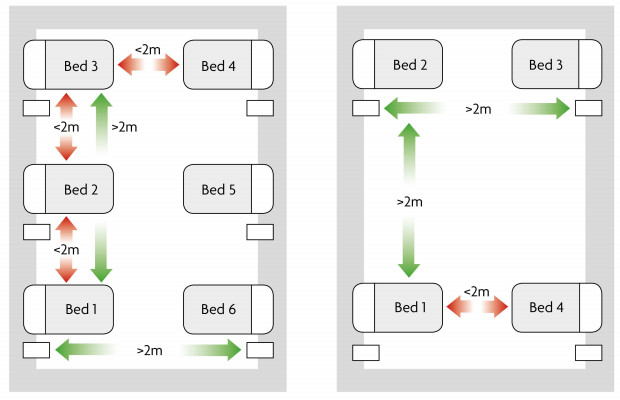
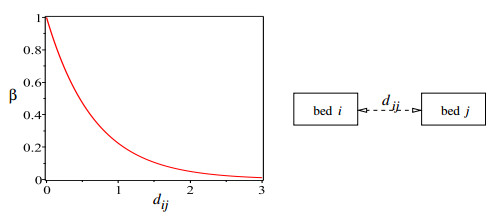
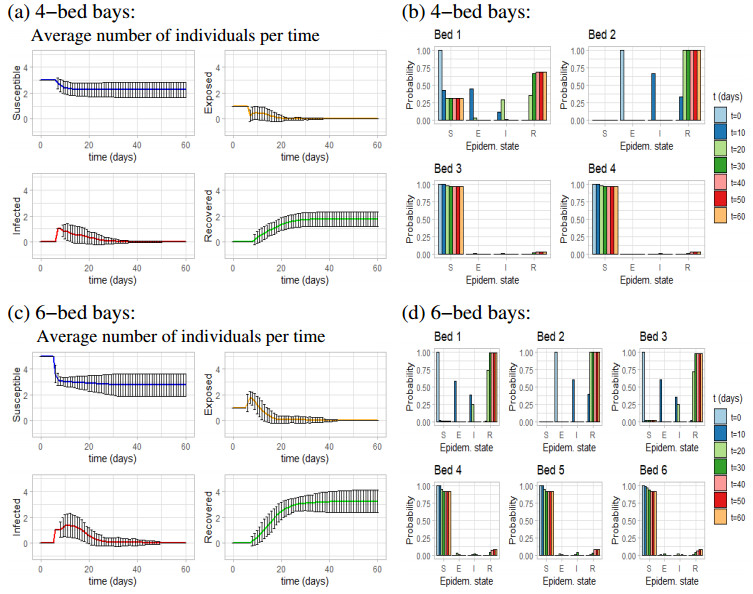
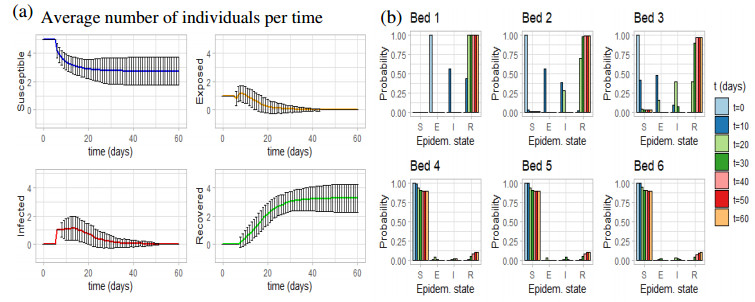
Figures(11) / Tables(1)

David Moreno Martos, Benjamin J. Parcell, Raluca Eftimie. Modelling the transmission of infectious diseases inside hospital bays: implications for COVID-19[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 8084-8104. doi: 10.3934/mbe.2020410







 DownLoad:
DownLoad: