Citation: Kwangjoong Kim, Wonhyung Choi. Local dynamics and coexistence of predator–prey model with directional dispersal of predator[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 6737-6755. doi: 10.3934/mbe.2020351

| [1] | M. Iida, M. Mimura, H. Ninomiya, Diffusion, cross-diffusion and competitive interaction, J. Math. Biol., 53 (2006), 617-641. |
| [2] |
W. Ko, K. Ryu, On a predator-prey system with cross diffusion representing the tendency of predators in the presence of prey species, J. Math. Anal. Appl., 341 (2008), 1133-1142. doi: 10.1016/j.jmaa.2007.11.018

|
| [3] |
T. Kadota, K. Kuto, Positive steady states for a prey-predator model with some nonlinear diffusion terms, J. Math. Anal. Appl., 323 (2006), 1387-1401. doi: 10.1016/j.jmaa.2005.11.065
|
| [4] | K. Kuto, A strongly coupled diffusion effect on the stationary solution set of a prey-predator model, Adv. Differential. Equ., 12 (2007), 145-172. |
| [5] | K. Kuto, Y. Yamada, Coexistence problem for a prey-predator model with density-dependent diffusion, Nonlinear Anal.-Theor., 71 (2009), e2223-e2232. |
| [6] | K. Kuto, Y. Yamada, Limiting characterization of stationary solutions for a prey-predator model with nonlinear diffusion of fractional type, Differ. Integral. Equ., 22 (2009), 725-752. |
| [7] |
Y. Lou, W. Ni, Diffusion vs cross-diffusion: an elliptic approach, J. Differ. Equations, 154 (1999), 157-190. doi: 10.1006/jdeq.1998.3559
|
| [8] |
K. Ryu, I. Ahn, Positive steady-states for two interacting species models with linear self-cross diffusions, Discrete Contin. Dyn-A, 9 (2003), 1049. doi: 10.3934/dcds.2003.9.1049
|
| [9] |
K. Ryu, I. Ahn, Coexistence theorem of steady states for nonlinear self-cross diffusion systems with competitive dynamics, J. Math. Anal. Appl., 283 (2003), 46-65. doi: 10.1016/S0022-247X(03)00162-8
|
| [10] |
N. Shigesada, K. Kawasaki, E. Teramoto, Spatial segregation of interacting species, J. Theor. Biol., 79 (1979), 83-99. doi: 10.1016/0022-5193(79)90258-3
|
| [11] | I. Averill, K. Lam, Y. Lou, The role of advection in a two-species competition model: a bifurcation approach, volume 245. American Mathematical Society, 2017. |
| [12] |
X. Chen, K. Lam, Y. Lou, Dynamics of a reaction-diffusion-advection model for two competing species, Discrete Contin. Dyn. S., 32 (2012), 3841-3859. doi: 10.3934/dcds.2012.32.3841
|
| [13] |
C. Cosner, Reaction-diffusion-advection models for the effects and evolution of dispersal, Discrete Contin. Dyn-A, 34 (2014), 1701-1745. doi: 10.3934/dcds.2014.34.1701
|
| [14] |
R. S. Cantrell, C. Cosner, Y. Lou, Movement toward better environments and the evolution of rapid diffusion, Math. Biosci., 204 (2006), 199-214. doi: 10.1016/j.mbs.2006.09.003
|
| [15] |
R. S. Cantrell, C. Cosner, Y. Lou, Advection-mediated coexistence of competing species, P. Roy. Soc. Edinb. A., 137 (2007), 497-518. doi: 10.1017/S0308210506000047
|
| [16] |
R. S. Cantrell, C. Cosner, Y. Lou, Approximating the ideal free distribution via reaction-diffusion- advection equations, J. Differ. Equations, 245 (2008), 3687-3703. doi: 10.1016/j.jde.2008.07.024
|
| [17] |
C. Cosner, Y. Lou, Does movement toward better environments always benefit a population?, J. Math. Anal. Appl., 277 (2003), 489-503. doi: 10.1016/S0022-247X(02)00575-9
|
| [18] |
K. Kuto, T. Tsujikawa, Limiting structure of steady-states to the lotka-volterra competition model with large diffusion and advection, J. Differ. Equations, 258 (2015), 1801-1858. doi: 10.1016/j.jde.2014.11.016
|
| [19] |
K.-Y. Lam, W.-M. Ni, Advection-mediated competition in general environments, J. Differ. Equations, 257 (2014), 3466-3500. doi: 10.1016/j.jde.2014.06.019
|
| [20] |
K.-Y. Lam, W. M. Ni, Limiting profiles of semilinear elliptic equations with large advection in population dynamics, Discrete Contin. Dyn. Syst., 28 (2010), 1051-1067. doi: 10.3934/dcds.2010.28.1051
|
| [21] |
E. Cho, Y.-J. Kim, Starvation driven diffusion as a survival strategy of biological organisms, B. Math. Biol., 75 (2013), 845-870. doi: 10.1007/s11538-013-9838-1
|
| [22] |
W. Choi, S. Baek, I. Ahn, Intraguild predation with evolutionary dispersal in a spatially heterogeneous environment, J. Math. Biol., 78 (2019), 2141-2169. doi: 10.1007/s00285-019-01336-5
|
| [23] |
W. Choi, I. Ahn, Strong competition model with non-uniform dispersal in a heterogeneous environment, Appl. Math. Lett., 88 (2019), 96-102. doi: 10.1016/j.aml.2018.08.014
|
| [24] |
W. Choi, I. Ahn, Non-uniform dispersal of logistic population models with free boundaries in a spatially heterogeneous environment, J. Math. Anal. Appl., 479 (2019), 283-314. doi: 10.1016/j.jmaa.2019.06.027
|
| [25] |
W. Choi, I. Ahn, Predator-prey interaction systems with non-uniform dispersal in a spatially heterogeneous environment, J. Math. Anal. Appl., 485 (2020), 123860. doi: 10.1016/j.jmaa.2020.123860
|
| [26] |
Y.-J. Kim, O. Kwon, F. Li, Evolution of dispersal toward fitness, B. Math. Biol., 75 (2013), 2474- 2498. doi: 10.1007/s11538-013-9904-8
|
| [27] |
Y.-J. Kim, O. Kwon, F. Li, Global asymptotic stability and the ideal free distribution in a starvation driven diffusion, J. Math. Biol., 68 (2014), 1341-1370. doi: 10.1007/s00285-013-0674-6
|
| [28] |
Y.-J. Kim, O. Kwon, Evolution of dispersal with starvation measure and coexistence, B. Math. Biol., 78 (2016), 254-279. doi: 10.1007/s11538-016-0142-8
|
| [29] |
W. Choi, I. Ahn, Effect of prey-taxis on predator's invasion in a spatially heterogeneous environment, Appl. Math. Lett., 98 (2019), 256-262. doi: 10.1016/j.aml.2019.06.021
|
| [30] |
S. Wu, J. Shi, B. Wu, Global existence of solutions and uniform persistence of a diffusive predator- prey model with prey-taxis, J. Differ. Equations, 260 (2016), 5847-5874. doi: 10.1016/j.jde.2015.12.024
|
| [31] |
H. Jin, Z. Wang, Global stability of prey-taxis systems, J. Differ. Equations, 262 (2017), 1257- 1290. doi: 10.1016/j.jde.2016.10.010
|
| [32] |
Y. Tao, Global existence of classical solutions to a predator-prey model with nonlinear prey-taxis, Nonlinear Anal.-Real., 11 (2010), 2056-2064. doi: 10.1016/j.nonrwa.2009.05.005
|
| [33] |
X. He, S. Zheng, Global boundedness of solutions in a reaction-diffusion system of predator-prey model with prey-taxis, Appl. Math. Lett., 49 (2015), 73-77. doi: 10.1016/j.aml.2015.04.017
|
| [34] |
C. Li, X. Wang, Y. Shao, Steady states of a predator-prey model with prey-taxis, Nonlinear Anal.- Theor., 97 (2014), 155-168. doi: 10.1016/j.na.2013.11.022
|
| [35] | P. A. Abrams, L. R. Ginzburg, The nature of predation: prey dependent, ratio dependent or neither?, Trends Ecol. Evol., 15 (2000), 337-341. |
| [36] |
R. Arditi, L. R. Ginzburg, Coupling in predator-prey dynamics: ratio-dependence, J. Theor. Biol., 139 (1989), 311-326. doi: 10.1016/S0022-5193(89)80211-5
|
| [37] |
C. Cosner, D. L. DeAngelis, J. S. Ault, D. B. Olson, Effects of spatial grouping on the functional response of predators, Theor. Popul. Biol., 56 (1999), 65-75. doi: 10.1006/tpbi.1999.1414
|
| [38] |
J. M. Culp, N. E. Glozier, G. J. Scrimgeour, Reduction of predation risk under the cover of darkness: avoidance responses of mayfly larvae to a benthic fish, Oecologia, 86 (1991), 163-169. doi: 10.1007/BF00317527
|
| [39] |
F. Mougeot, V. Bretagnolle, Predation risk and moonlight avoidance in nocturnal seabirds, J. Avian. Biol., 31 (2000), 376-386. doi: 10.1034/j.1600-048X.2000.310314.x
|
| [40] | T. Caro, Antipredator defenses in birds and mammals, University of Chicago Press, 2005. |
| [41] | H. B. Cott, Adaptive coloration in animals, 1940. |
| [42] |
J. M. Hemmi, Predator avoidance in fiddler crabs: 1. escape decisions in relation to the risk of predation, Anim. Behav., 69 (2005), 603-614. doi: 10.1016/j.anbehav.2004.06.018
|
| [43] | W. J. Bell, Searching behaviour: the behavioural ecology of finding resources, Springer Science & Business Media, 2012. |
| [44] |
S. Benhamou, Spatial memory and searching efficiency, Anim. Behav., 47 (1994), 1423-1433. doi: 10.1006/anbe.1994.1189
|
| [45] |
S. Benhamou, Bicoordinate navigation based on non-orthogonal gradient fields, J. Theo. Biol., 225 (2003), 235-239. doi: 10.1016/S0022-5193(03)00242-X
|
| [46] | W. F. Fagan, M. A. Lewis, M. Auger-Meth ′ e, T. Avgar, S. Benhamou, G. Breed, et al., Spatial ′ memory and animal movement, Ecol. Lett., 16 (2013), 1316-1329. |
| [47] |
S. M. Flaxman, Y. Lou, Tracking prey or tracking the prey's resource? mechanisms of movement and optimal habitat selection by predators, J. Theor. Biol., 256 (2009), 187-200. doi: 10.1016/j.jtbi.2008.09.024
|
| [48] |
S. M. Flaxman, Y. Lou, F. G. Meyer, Evolutionary ecology of movement by predators and prey, Theor. Ecol., 4 (2011), 255-267. doi: 10.1007/s12080-011-0120-6
|
| [49] | A. M. Kittle, M. Anderson, T. Avgar, J. A. Baker, G. S. Brown, J. Hagens, et al., Landscape-level wolf space use is correlated with prey abundance, ease of mobility, and the distribution of prey habitat, Ecosphere, 8 (2017), e01783. |
| [50] | H. Amann, Nonhomogeneous linear and quasilinear elliptic and parabolic boundary value problems, In Function spaces, differential operators and nonlinear analysis, pages 9-126. Springer, 1993. |
| [51] | R. S. Cantrell, C. Cosner, Spatial ecology via reaction-diffusion equations, John Wiley & Sons, 2004. |
| [52] |
L. J. S. Allen, B. M. Bolker, Y. Lou, A. L. Nevai, Asymptotic profiles of the steady states for an sis epidemic reaction-diffusion model, Discrete Contin. Dyn. Syst. Ser. A, 21 (2008), 1. doi: 10.3934/dcds.2008.21.1
|
| [53] |
X. He, W.-M. Ni, Global dynamics of the lotka-volterra competition-diffusion system: Diffusion and spatial heterogeneity i, Commun. Pur. Appl. Math., 69 (2016), 981-1014. doi: 10.1002/cpa.21596
|
| [54] |
E. N. Dancer, On the indices of fixed points of mappings in cones and applications, J. Math. Anal. Appl., 91 (1983), 131-151. doi: 10.1016/0022-247X(83)90098-7
|
| [55] |
L. Li, Coexistence theorems of steady states for predator-prey interacting systems, T. Am. Math. Soc., 305 (1988), 143-166. doi: 10.1090/S0002-9947-1988-0920151-1
|
| [56] | M. Wang, Z. Li, Q. Ye, Existence of positive solutions for semilinear elliptic system, In Qualitative aspects and applications of nonlinear evolution equations, 1991. |
| [57] |
K. Ryu, I. Ahn, Positive solutions for ratio-dependent predator-prey interaction systems, J. Differ. Equations, 218 (2005), 117-135. doi: 10.1016/j.jde.2005.06.020
|
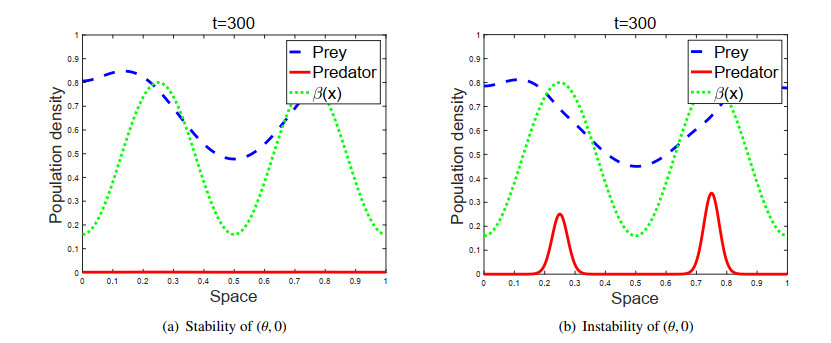
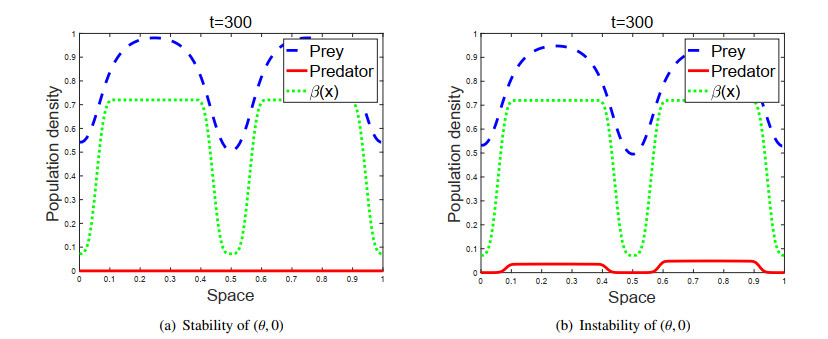
Figures(2)

Kwangjoong Kim, Wonhyung Choi. Local dynamics and coexistence of predator–prey model with directional dispersal of predator[J]. Mathematical Biosciences and Engineering, 2020, 17(6): 6737-6755. doi: 10.3934/mbe.2020351







 DownLoad:
DownLoad: