Citation: Fumin Zhang, Zhipeng Qiu, Balian Zhong, Tao Feng, Aijun Huang. Modeling Citrus Huanglongbing transmission within an orchard and its optimal control[J]. Mathematical Biosciences and Engineering, 2020, 17(3): 2048-2069. doi: 10.3934/mbe.2020109

| [1] | J. M. Bové, Huanglongbing: a destructive, newly-emerging, century-old disease of citrus, J. Plant. Pathol., 88 (2006), 7-37. |
| [2] | R. A. Taylor, E. A. Mordecai, C. A. Gilligan, J. R. Rohr, L. R. Johnson, Mathematical models are a powerful method to understand and control the spread of Huanglongbing, Peer J., 19 (2016), 2642. |
| [3] | J. M. Bové, M. E. Rogers, Huanglongbing-control workshop: summary, Acta. Hort., 1065 (2015), 869-889. |
| [4] | A. J. Ayres, J. B. Jr, J. M. Bové, The experience with Huanglongbing management in Brazil, Acta. Hort., 1065 (2015), 55-61. |
| [5] | K. Chen, C. Li, Survey of citrus Huanglongbing research, J. Zhejiang Agr. Sci., 56 (2015), 1048-1050. (in Chinese) |
| [6] | C. Chiyaka, B. H. Singer, S. E. Halbert, J. G. M. Jr, A. H. C. van Bruggen, Modeling Huanglongbing transmission within a citrus tree, Proc. Natl. Acad. Sci. USA, 109 (2012), 12213-12218. |
| [7] | M. S. Chan, M. J. Jeger, An analytical model of plant virus disease dynamics with roguing and replanting, J. Appl. Ecol., 31 (1994), 413-427. |
| [8] | M. J. Jeger, L. V. Madden, F. van den Bosch, The effect of transmission route on plant virus epidemic development and disease control, J. Theor. Biol., 258 (2009), 198-207. |
| [9] | M. S. Sisterson, Effects of insect-vector preference for healthy or infected plants on pathogen spread: Insights from a model, J. Econ. Entomol., 101 (2008), 1-8. |
| [10] | X. Z. Meng, Z. Q. Li, The dynamics of plant disease models with continuous and impulsive cultural control strategies, J. Theor. Biol., 266 (2010), 29-40. |
| [11] | X. Z. Meng, Z. T. Song, L. S. Chen, A new mathematical model for optimal control strategies of integrated pest management, J. Biol. Syst., 15, (2007), 219-234. |
| [12] | W. C. Zhao, J. Li, T. Q. Zhang, X. Z. Meng, T. H. Zhang, Persistence and ergodicity of plant disease model with Markov conversion and impulsive toxicant input, Commun. Nonlinear Sci., 48 (2017), 70-84. |
| [13] | L. J. Xia, S. J. Gao, Q. Zou, J. P. Wang, Analysis of a nonautonomous plant disease model with latent period, Appl. Math. Comput., 223 (2013), 147-159. |
| [14] | S. J. Gao, L. J. Xia, J. L. Wang, Z. J. Zhang, Modelling the effects of cross protection control in plant disease with seasonality, Int. J. Biomath., 10 (2017), 1-24. |
| [15] | S. J. Gao, D. Yu, X. Z. Meng, F. M. Zhang, Global dynamics of a stage-structured Huanglongbing model with time delay, Chaos. Soliton. Fract., 117 (2018), 60-67. |
| [16] | X. S. Zhang, J. Holt, J. Colvin, Mathematical models of host plant infection by helper-dependent virus complexes: why are helper viruses always avirulent, Phytopathology, 90 (2000), 85-93. |
| [17] | A. U. Awan, M. Ozair, Stability analysis of pine wilt disease model by periodic use of insecticides, J. Biol. Dynam., 10 (2016), 506-524. |
| [18] | M. A. Khan, R. Khan, Y. Khan, S. Islam, A mathematical analysis of Pine Wilt disease with variable population size and optimal control strategies, Chaos Soliton Fract., 108 (2018), 205-217. |
| [19] | K. Jacobsen, J. Stupiansky, S. S. Pilyugin, Mathematical modeling of citrus groves infected by Huanglongbing, Math. Biosci. Eng., 10 (2013), 705-728. |
| [20] | X. S. Zhang, J. Holt, J. Colvin, Synergism between plant viruses: a mathematical analysis of the epidemiological implications, Plant Pathol., 50 (2001), 732-746. |
| [21] | K. S. Lee, D. Kim, Global dynamics of a pine wilt disease transmission model with nonlinear incidence rates, Appl. Math. Model, 37 (2013), 4561-4569. |
| [22] | M. Ozair, X. Y. Shi, T. Hussain, Control measures of pine wilt disease, Comp. Appl. Math., 35 (2016), 519-531. |
| [23] | M. A. Khan, K. Ali, E. Bonyah, K. O. Okosun, S. Islam, A. Khan, Mathematical modeling and stability analysis of Pine Wilt Disease with optimal control, Sci. Rep., 7 (2017), 3115. |
| [24] | M. A. Khan, R. Khan, Y. Khan, S. Islam, A mathematical analysis of Pine Wilt disease with variable population size and optimal control strategies, Chaos Soliton Fract., 108 (2018), 205-217. |
| [25] | J. A. Lee, S. E. Halbert, W. O. Dawson, C. J. Robertson, J. E. Keesling, B. H. Singer, Asymptomatic spread of Huanglongbing and implications for disease control, Proc. Natl. Acad. Sci. USA, 112 (2015), 7605-7610. |
| [26] | R. G. dA. Vilamiu, S. Ternes, G. A. Braga, F. F. Laranjeira, A model for Huanglongbing spread between citrus plants including delay times and human intervention, AIP Conf. Proc., 1479 (2012), 2315-2319. |
| [27] | Wikipedia, Available from: https://en.wikipedia.org/wiki/Citrus_greening-disease. |
| [28] | M. X. Deng, Forming process and basis and technological points of the theory emphasis on control citrus psylla for integrated control Huanglongbing, Chin. Agric. Sci. Bull., 25 (2009), 358-363. |
| [29] | O. Diekmann, J. A. P. Heesterbeek, J. A. J. Metz, On the definition and the computation of the basic reproduction ratio $\mathscr{R}_0$ in models for infectious diseases in heterogeneous populations, J. Math. Biol., 28 (1990), 365-382. |
| [30] | P. Dreessche, J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29-48. |
| [31] | X. Q. Zhao, Dynamical Systems in Population Biology (Second Edition), Springer-Verlag, New York, 2017. |
| [32] | H. L. Smith, Monotone Dynamical System: An Introduction to the Theory of Competitive and Cooperative Systems, American Mathematical Society, Providence, 1995. |
| [33] | C. Castillo-Chavez, H. Thieme, Asymptotically autonomous epidemic models, Math. Popul. Dyn.: Anal. Heterog., 1 (1995), 33-50. |
| [34] | H. R. Thieme, Persistence under relaxed point-dissipativity with an application to an epidemic model, SIAM J. Math. Anal., 24 (1993), 407-435. |
| [35] | R. Gamkrelidze, L. S. Pontrjagin, V. G. Boltjanskij, The Mathematical Theory of Optimal Processes, Macmillan Company, 1964. |
| [36] | S. M. Blower, H. Dowlatabadi, Sensitivity and uncertainty analysis of complex models of disease transmission: An HIV model, as an example, Int. Stat. Rev., 62 (1994), 229-243. |
| [37] | J. C. Helton, J. D. Johnson, C. J. Sallaberry, C. B. Storlie, Survey of sampling-based methods for uncertainty and sensitivity analysis, Reliab. Eng. Syst. Saf., 91 (2006), 1175-1209. |
| [38] | S. Marino, I. B. Hogue, C. J. Ray, D. E. Kirschner, A methodology for performing global uncertainty and sensitivity analysis in systems biology, J. Theor. Biol., 254 (2008), 178-196. |
| [39] | M. A. Sanchez, S. M. Blower, Uncertainty and sensitivity analysis of the basic reproductive rate: tuberculosis as an example, Am. J. Epidemiol., 145 (1997), 1127-1137. |
| [40] | S. Lenhart, J. T. Workman, Optimal Control Applied to Biological Models, Chapman and Hall/CRC, Boca Raton, FL, 2007. |
| [41] | X. F. Yan, Y. Zou, Optimal and sub-optimal quarantine and isolation control in SARS epidemics, Math. Comput. Model., 47 (2008), 235-245. |
| [42] | F. B. Agusto, M. A. Khan, Optimal control strategies for dengue transmission in pakistan, Math. Biosci., 305 (2018), 102-121. |
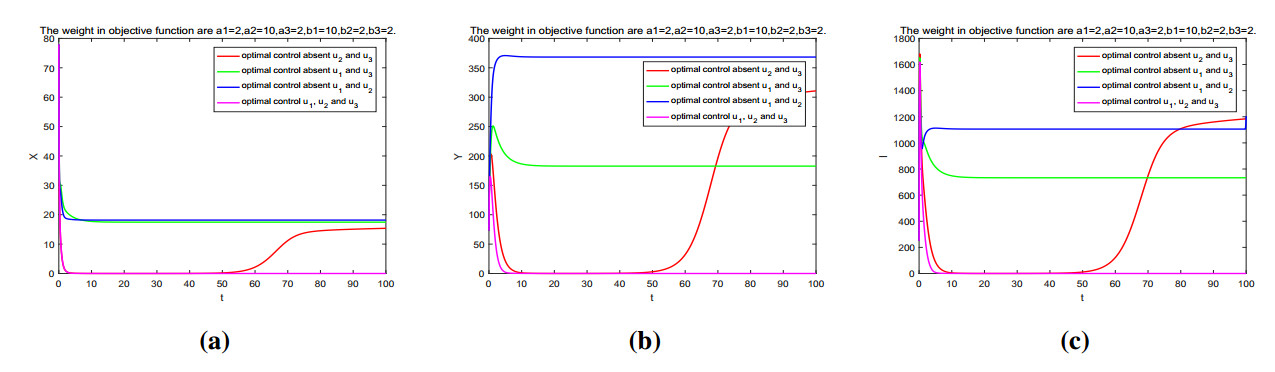
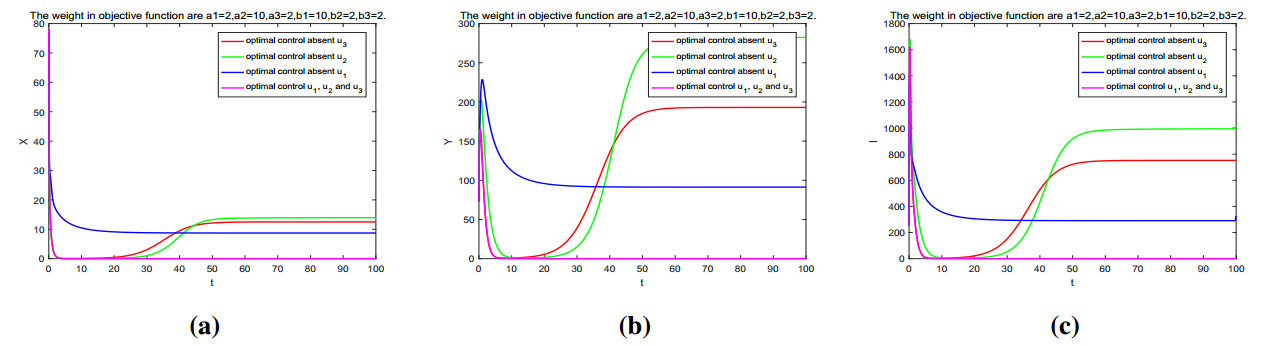
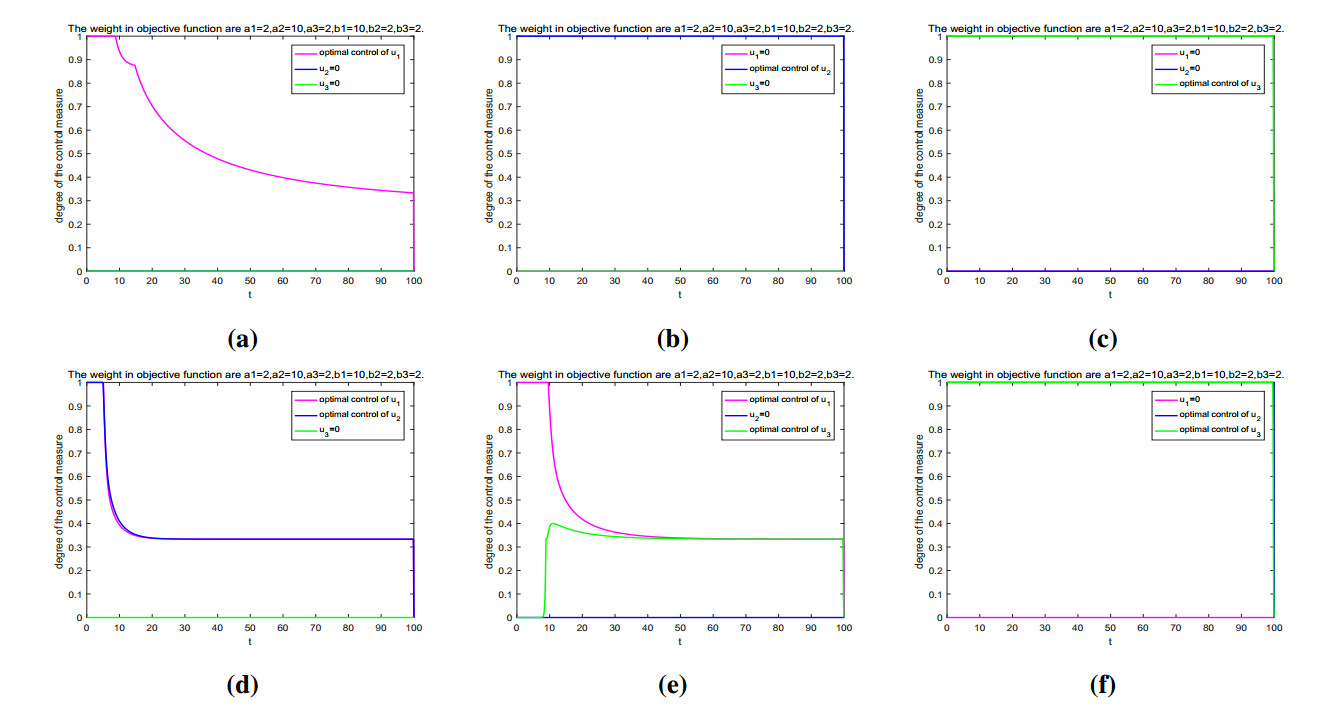
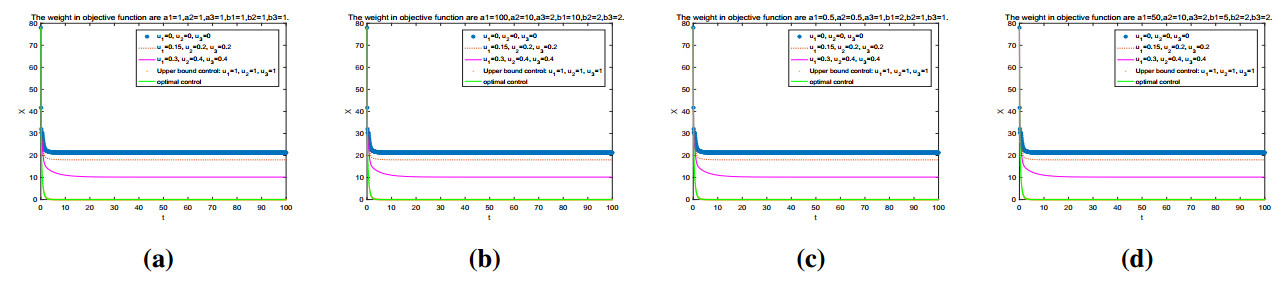
Figures(11) / Tables(2)

Fumin Zhang, Zhipeng Qiu, Balian Zhong, Tao Feng, Aijun Huang. Modeling Citrus Huanglongbing transmission within an orchard and its optimal control[J]. Mathematical Biosciences and Engineering, 2020, 17(3): 2048-2069. doi: 10.3934/mbe.2020109







 DownLoad:
DownLoad: