Citation: Tianqi Song, Chuncheng Wang, Boping Tian. Mathematical models for within-host competition of malaria parasites[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 6623-6653. doi: 10.3934/mbe.2019330

| [1] | World malaria report, World Health Organization (2017). |
| [2] | L. C. Pollitt, S. Huijben, D. G. Sim, et al., Rapid response to selection, competitive release and increased transmission potential of artesunate-selected plasmodium chabaudi malaria parasites, PLoS Pathog., 10 (2014), e1004019. |
| [3] | E. Hansen, R. J. Woods and A. F. Read, How to use a chemotherapeutic agent when resistance to it threatens the patient, PloS Biol., 15 (2017), e2001110. |
| [4] | A. R. Wargo, S. Huijben, J. C. D. Roode, et al., Competitive release and facilitation of drug-resistant parasites after therapeutic chemotherapy in a rodent malaria model, Proc. Natl. Acad. Sci. USA, 104 (2007), 19914–19919. |
| [5] | D. I. Andersson and D. Hughes, Antibiotic resistance and its cost: is it possible to reverse resis-tance? Nat. Rev. Microbiol., 8 (2010), 260–271. |
| [6] | I. Hastings, A model for the origins and spread of drug-resistant malaria, Parasitology, 115 (1997), 133–141. |
| [7] | M. Bushman, L. Morton, N. Duah, et al., Within-host competition and drug resistance in the human malaria parasite plasmodium falciparum, Proc. R. Soc. B, 283 (2016), 20153038. |
| [8] | A. F. Read and H. Silvie, Evolutionary biology and the avoidance of antimicrobial resistance, Evol. Appl., 2 (2009), 40–51. |
| [9] | R. Ataide, E. A. Ashley, R. Powell, et al., Host immunity to plasmodium falciparum and the assessment of emerging artemisinin resistance in a multinational cohort, Proc. Natl. Acad. Sci. USA, 114 (2017), 3515–3520. |
| [10] | A. Handel and B. M. Elevin, Exploring the role of the immune response in preventing antibiotic resistance, J. Theor. Biol., 256 (2009), 655–662. |
| [11] | K. Drlica and X. Zhao, Mutant selection window hypothesis updated, Clin. Infect. Dis., 44 (2007), 681–688. |
| [12] | E. Gjini and P. H. Brito, Integrating antimicrobial therapy with host immunity to fight drug-resistant infections: classical vs. adaptive treatment, PLoS Comput. Biol., 12 (2016), e1004857. |
| [13] | M. J. Mackinnon, Drug resistance models for malaria, Acta Trop., 94 (2005), 207–217. |
| [14] | J. M. Tchuenche, C. Chiyaka, D. Chan, et al., A mathematical model for antimalarial drug resis-tance, Math. Med. Biol., 28 (2011), 335–355. |
| [15] | J. Hansen and T. Day, Coinfection and the evolution of drug resistance, J. Evol. Biol., 27 (2015), 2595–2604. |
| [16] | C. Chiyaka, W. Garira and S. Dube, Modelling immune response and drug therapy in human malaria infection, Comput. Math. Method Med., 9 (2008), 143–163. |
| [17] | Y. Li, S. Ruan and D. Xiao, The within-host dynamics of malaria infection with immune response., Math. Biosci. Eng., 8 (2011), 999–1018. |
| [18] | M. Bushman, R. Antia, V. Udhayakumar, et al., Within-host competition can delay evolution of drug resistance in malaria, PLoS Biol., 16 (2018), e2005712. |
| [19] | L. Mathieu and B. Sebastian, A combined within-host and between-hosts modelling framework for the evolution of resistance to antimalarial drugs, J. R. Soc. Interface, 13 (2016), 20160148. |
| [20] | C. Hetzel and R. M. Anderson, The within-host cellular dynamics of bloodstage malaria: theoret-ical and experimental studies, Parasitology, 113 (1996), 25–38. |
| [21] | R. M. Anderson, R. M. May and S. Gupta, Non-linear phenomena in host-parasite interactions, Parasitology, 99 (1989), S59–S79. |
| [22] | C. E. Cressler, W. A. Nelson, T. Day, et al., Disentangling the interaction among host resources, the immune system and pathogens, Ecol. Lett., 17 (2014), 284–293. |
| [23] | P. B. Greenspoon, S. Banton and N. Mideo, Immune system handling time may alter the outcome of competition between pathogens and the immune system, J. Theor. Biol., 447 (2018), 25–31. |
| [24] | P. Dreessche and J. Watmough, Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. |
| [25] | D. Xiao and W. H. Bossert, An intra-host mathematical model on interaction between hiv and malaria, Bull. Math. Biol., 72 (2010), 1892–1911. |
| [26] | S. Ruan and S. K. W. Gail, Bifurcation analysis of a chemostat model with a distribued delay, J. Math. Anal. Appl., 204 (1996), 786–812. |
| [27] | A. Saltelli, P. Annoni, I. Azzini, et al., Variance based sensitivity analysis of model output. design and estimator for the total sensitivity index, Comput. Phys. Commun., 181 (2010), 259–270. |
| [28] | I. M. Sobol, Sensitivity estimates for nonlinear mathematical models, in Mathematical Modeling and Computational Experiment, 2010. |
| [29] | N. Wale, D. G. Sim, M. J. Jones, et al., Resource limitation prevents the emergence of drug resis-tance by intensifying within-host competition, Proc. Natl. Acad. Sci. USA, 114 (2017), 13774. |
| [30] | A. F. Read, T. Day and S. Huijben, The evolution of drug resistance and the curious orthodoxy of aggressive chemotherapy, Proc. Natl. Acad. Sci. USA, 108 (2011), 10871–10877. |
| [31] | J. Stark, C. Chan and A. J. George, Oscillations in the immune system., Immunol. Rev., 216 (2007), 213–231. |
| [32] | B. Hellriegel, Modelling the immune response to malaria with ecological concepts: short-term behavior against long-term equilibrium, Proc. R. Soc. B-Biol. Sci., 250 (1992), 249–256. |
| [33] | P. Ankomah and B. R. Levin, Exploring the collaboration between antibiotics and the immune response in the treatment of acute, self-limiting infections, Proc. Natl. Acad. Sci. USA, 111 (2014), 8331–8. |
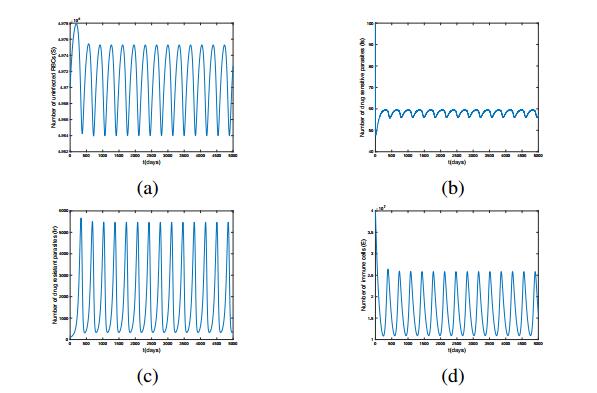
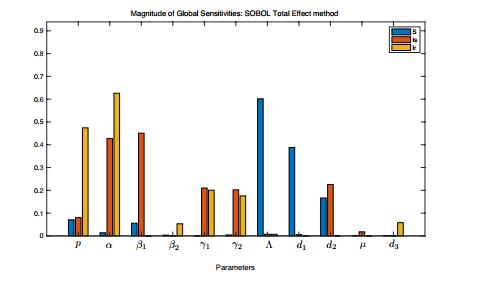
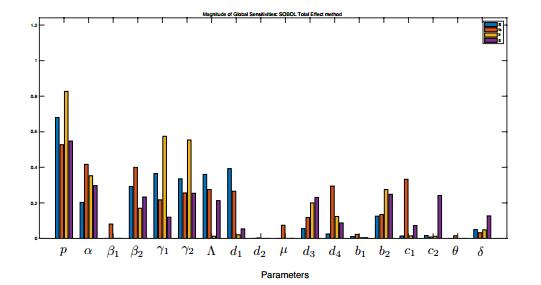
Figures(11) / Tables(1)

Tianqi Song, Chuncheng Wang, Boping Tian. Mathematical models for within-host competition of malaria parasites[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 6623-6653. doi: 10.3934/mbe.2019330







 DownLoad:
DownLoad: