Citation: Huihao Wang, Kuan Wang, Zhen Deng, Xiaofei Li, Yi-Xian Qin, Hongsheng Zhan, Wenxin Niu. Effects of facet joint degeneration on stress alterations in cervical spine C5–C6: A finite element analysis[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 7447-7457. doi: 10.3934/mbe.2019373

| [1] | N. Bogduk, The anatomy and pathophysiology of neck pain, Phys. Med. Rehabil. Clin. N. Am., 22 (2011), 367–382. |
| [2] | N. Yoganandan, S. Kumaresan and F. A. Pintar, Biomechanics of the cervical spine Part 2. Cervical spine soft tissue responses and biomechanical modeling, Clin. Biomech. (Bristol, Avon), 16 (2001), 1–27. |
| [3] | M. Richter, H. J. Wilke, P. Kluger, et al., Load-displacement properties of the normal and injured lower cervical spine in vitro, Eur. Spine J., 9 (2000), 104–108. |
| [4] | R. A. Hartman, R. E. Tisherman, C. Wang, et al., Mechanical role of the posterior column components in the cervical spine, Eur. Spine J., 25 (2016), 2129–2138. |
| [5] | N. Yoganandan, S. Kumaresan and F. A. Pintar, Geometric and mechanical properties of human cervical spine ligaments. J. Biomech. Eng., 122 (2000), 623–629. |
| [6] | S. F. Mattucci, J. A. Moulton, N. Chandrashekar, et al., Strain rate dependent properties of younger human cervical spine ligaments, J. Mech. Behav. Biomed. Mater., 10 (2012), 216–226. |
| [7] | D. Steilen, R. Hauser, B. Woldin, et al., Chronic neck pain: Making the connection between capsular ligament laxity and cervical instability, Open Orthop. J., 8 (2014), 326–345. |
| [8] | T. Iida, K. Abumi, Y. Kotani, et al., Effects of aging and spinal degeneration on mechanical properties of lumbar supraspinous and interspinous ligaments, Spine J., 2 (2002), 95–100. |
| [9] | K. P. Quinn, K. E. Lee, C. C. Ahaghotu, et al., Structural changes in the cervical facet capsular ligament: Potential contributions to pain following subfailure loading, Stapp Car Crash J., 51 (2007), 169–187. |
| [10] | G. P. Pal, R. V. Routal and S. K. Saggu, The orientation of the articular facets of the zygapophyseal joints at the cervical and upper thoracic region, J. Anat., 198 (2001), 431–441. |
| [11] | X. Rong, Z. Liu, B. Wang, et al., Relationship between facet tropism and facet joint degeneration in the sub-axial cervical spine, BMC Musculoskelet. Disord., 18 (2017), 86. |
| [12] | H. Zeng, D. Zou and J. Wu, Helical CT three-dimensional reconstruction for inferior cervical zygapophyseal joint and its clinic meaning, Chi. J. Spine Spinal Cord, 1 (2012), 59–64. |
| [13] | R. C. Decker, Surgical treatment and outcomes of cervical radiculopathy, Phys. Med. Rehabil. Clin. N. Am., 22 (2011) 179–191. |
| [14] | Z. Cai, Z. Li, J. Dong, et al., A study on protective performance of bullet-proof helmet under impact loading, J. Vibroeng, 18 (2016), 2495–2507. |
| [15] | T. Xie, J. Qian, Y. Lu, et al., Biomechanical comparison of laminectomy, hemilaminectomy and a new minimally invasive approach in the surgical treatment of multilevel cervical intradural tumour: a finite element analysis, Eur. Spine J., 22 (2013), 2719–2730. |
| [16] | B. D. Stemper, N. Yoganandan and F. A. Pintar, Effects of abnormal posture on capsular ligament elongations in a computational model subjected to whiplash loading, J. Biomech., 38 (2005), 1313–1323. |
| [17] | J. D. Hoppenfeld, Cervical facet arthropathy and occipital neuralgia: headache culprits, Curr. Pain Headache Rep., 14 (2010), 418–423. |
| [18] | K. Wang, H. Wang, Z. Deng, et al., Cervical traction therapy with and without neck support: A finite element analysis, Musculoskelet. Sci. Pract., 28 (2017), 1–9. |
| [19] | Z. B. Friedenberg and W. T. Miller, Degenerative disc disease of the cervical spine, J. Bone Joint Surg. Am., 45 (1963), 1171–1178. |
| [20] | Z. Wang, H. Zhao, J. M. Liu, et al., Resection or degeneration of uncovertebral joints altered the segmental kinematics and load-sharing pattern of subaxial cervical spine: a biomechanical investigation using a C2–T1 finite element model, J. Biomech., 49 (2016), 2854–2862. |
| [21] | N. Kallemeyn, A. Gandhi, S. Kode, et al., Validation of a C2–C7 cervical spine finite element model using specimen-specific flexibility data, Med. Eng. Phys., 32 (2010), 482–489. |
| [22] | M. B. Panzer and D. S. Cronin, C4–C5 segment finite element model development, validation, and load-sharing investigation, J. Biomech., 42 (2009), 480–490. |
| [23] | S. F. Mattucci and D. S. Cronin, A method to characterize average cervical spine ligament response based on raw data sets for implementation into injury biomechanics models, J. Mech. Behav. Biomed. Mater., 41 (2015), 251–260. |
| [24] | S. H. Lee, Y. J. Im, K. T. Kim, et al., Comparison of cervical spine biomechanics after fixed- and mobile-core artificial disc replacement: a finite element analysis, Spine (Phila Pa 1976), 36 (2011), 700–708. |
| [25] | Y. Wu, J. Wu, J. Guan, et al., Study of double-level degeneration of lower lumbar spines by finite element model, World Neurosurg., 86 (2015), 294–299. |
| [26] | Z. Mo, Y. Zhao, C. Du, et al., Does location of rotation center in artificial disc affect cervical biomechanics?, Spine (Phila Pa 1976), 40 (2015), 469–475. |
| [27] | K. Wang, Z. Deng, H. Wang, et al., Influence of variations in stiffness of cervical ligaments on C5–C6 segment, J. Mech. Behav. Biomed. Mater., 72 (2017), 129–137. |
| [28] | S. A. O'Leary, J. M. Link, E. O. Klineberg, et al., Characterization of facet joint cartilage properties in the human and interspecies comparisons. Acta. Biomater., 54 (2017), 367–376. |
| [29] | J. Dvorak, J. A. Antinnes, M. Panjabi, et al., Age and gender related normal motion of the cervical spine, Spine (Phila Pa 1976), 17 (1992), 393–398. |
| [30] | F. Heuer, H. Schmidt, Z. Klezl, et al., Stepwise reduction of functional spinal structures increase range of motion and change lordosis angle, J. Biomech., 40 (2007), 271–280. |
| [31] | K. P. Quinn, K. E. Lee, C. C. Ahaghotu, et al., Structural changes in the cervical facet capsular ligament: Potential contributions to pain following subfailure loading, Stapp Car Crash J., 51 (2007), 169–187. |
| [32] | P. D. Leahy and C. M. Puttlitz, The effects of ligamentous injury in the human lower cervical spine, J. Biomech., 45 (2012), 2668–2672. |
| [33] | Y. Kotani, P. S. McNulty, K. Abumi, et al., The role of anteromedial foraminotomy and the uncovertebral joints in the stability of the cervical spine, a biomechanical study, Spine (Phila Pa 1976), 23 (1998), 1559–1565. |
| [34] | Z. Cai, Z. Li, L. Wang, et al., A three-dimensional finite element modelling of human chest injury following front or side impact loading, J. Vibroeng., 18 (2016), 539–550. |
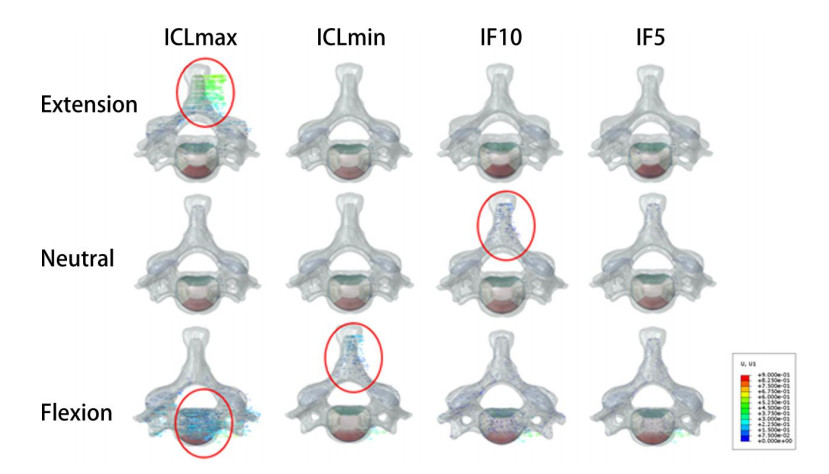

Figures(6) / Tables(1)

Huihao Wang, Kuan Wang, Zhen Deng, Xiaofei Li, Yi-Xian Qin, Hongsheng Zhan, Wenxin Niu. Effects of facet joint degeneration on stress alterations in cervical spine C5–C6: A finite element analysis[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 7447-7457. doi: 10.3934/mbe.2019373







 DownLoad:
DownLoad: