Citation: Sarah Treibert, Helmut Brunner, Matthias Ehrhardt. Compartment models for vaccine effectiveness and non-specific effects for Tuberculosis[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 7250-7298. doi: 10.3934/mbe.2019364

| [1] | B. Brodhun, D. Altman, B. Hauer, et al., Bericht zur Epidemiologie der Tuberkulose in Deutschland für 2017 (Report on the epidemiology of tuberculosis in Germany for 2017) (in german), Robert Koch-Institut, Berlin 2018. |
| [2] | K. Bozorgmehr, C. Stock, B. Joggerst, et al., Tuberculosis screening in asylum seekers in Germany: a need for better data, The Lancet, 3 (2018), Pe359–e361. |
| [3] | K. Bozorgmehr, S. Preussler, U. Wagner, et al., Using country of origin to inform targeted tuberculosis screening in asylum seekers: a modelling study of screening data in a German federal state, 2002-2015, BMC Infect. Diseases, 19 (2019), 304. |
| [4] | R. Diel, S. Rüsch-Gerdes and S. Niemann, Molecular Epidemiology of Tuberculosis among Immigrants in Hamburg, Germany, J. Clin. Microbiol., 42 (2004), 2952–2960. |
| [5] | H. Guoand J. Wu, Persistenthigh incidence of tuberculosisamong immigrants in alow-incidence country: impact of immigrants with early or late latency, Math. Biosci. Eng., 8 (2011), 695–709. |
| [6] | J. Zhang, Y. Li and X. Zhang, Mathematical modeling of tuberculosis data of China, J. Theor. Biol., 365 (2015), 159–163. |
| [7] | Focus, Tuberkulosefälle in Deutschland nehmen wieder zu – vor allem in den Großstädten (Tuberculosis cases in Germany on the rise again – especially in large cities) (in german), March 23, 2017, |
| [8] | R. W. Aldridge, D. Zenner, P. J. White, et al., Tuberculosis in migrants moving from high-incidence to low-incidence countries: a population-based cohort study of 519 955 migrants screened before entry to England, Wales, and Northern Ireland, The Lancet, October 11, 2016. |
| [9] | N. A. Menzies, A. N. Hill, T. Cohen, et al., The impact of migration on tuberculosis in the United States, Int. J. Tuberc. Lung Dis., 22 (2018), 1392–1403. |
| [10] | D. P. Moualeu, S. Röblitz, R. Ehrig, et al., Parameter Identification for a Tuberculosis Model in Cameroon, PLoS ONE, 10 (2015), e0120607. |
| [11] | D. P. Moualeu, A. N. Yakam, S. Bowong, et al., Analysis of a tuberculosis model with undetected and lost-sight cases, Commun. Nonlin. Sci. Numer. Simul., 41 (2016), 48–63. |
| [12] | M. Pareek, C. Greenaway, T. Noori, et al., The impact of migration on tuberculosis epidemiology and control in high-income countries: a review, BMC Med., 14 (2016), 48. |
| [13] | R. P. Sigdel and C. C. McCluskey, Global stability for an SEI model of infectious disease with immigration, Appl. Math. Comput., 243 (2014), 684–689. |
| [14] | Z. White, J. Painter, P. Douglas, et al., Immigrant Arrival and Tuberculosis among Large Immigrant- and Refugee-Receiving Countries, 2005–2009, Tuberc. Res. Treat., 2017 (2017), Article 8567893. |
| [15] | Y. Zhou and H. Cao, Discrete tuberculosis models and their application, in: S. Sivaloganathan (ed.), New Perspectives in Mathematical Biology, Fields Institute Communications 57, 2010. |
| [16] | R. Loddenkemper, J. F. Murray, C. Gradmann, et al., History of tuberculosis, Chapter 2 in: G.B. Migliori, G. Bothamley, R. Duarte, A. Rendon (eds.) Tuberculosis, ERS Monograph 82, 2018. |
| [17] | N. E. Aronson, M. Santosham, G. W. Comstock, et al., Long-term efficacy of BCG vaccine in American indians and Alaska natives, JAMA, 291 (2004), 2086–2091. |
| [18] | J. P. Higgins, K. Soares-Weiser, J. A. Lopez-Lopez, et al., Association of BCG, DTP, and measles containing vaccines with childhood mortality: systematic review, BMJ, 355 (2016), i5170. |
| [19] | P. Aaby, T. R. Kollmann and C. S. Benn, Nonspecific effects of neonatal and infant vaccination: public-health, immunological and conceptual challenges, Nat. Immunol., 15 (2014), 895–899. |
| [20] | S. Biering-Sørensen, K. J. Jensen, I. Monterio, et al., Rapid protective effects of early BCG on neonatal mortality among low birth weight boys: Observations from randomized trials, J. Infect. Dis., 217 (2018), 759–766. |
| [21] | V. Nankabirwa, J. K. Tumwine, O. Namugga, et al., Early versus late BCG vaccination in HIV-1-exposed infants in Uganda: study protocol for a randomized controlled trial, Trials, 18 (2017), 152–169. |
| [22] | M. G. Netea, L. A. Joosten, E. Latz, et al., Trained immunity: A program of innate immune memory in health and disease, Science, 352 (2016), aaf1098. |
| [23] | M. G. Netea, J. Quintin and J. van der Meer, Trained Immunity: A memory for innate host defense, Cell Press, Cell Host and Microbe, 2011. |
| [24] | M. G. Netea, Training innate immunity: The changing concept of immunological memory in innate host defence, Eur. J. Clin. Invest., 43 (2013), 881–884. |
| [25] | F. Shann, The non-specific effects of vaccines, Arch. Dis. Child., 95 (2010), 662–667. |
| [26] | C. B. Wilson, Applying contemporary immunology to elucidate heterologous effects of infant vaccines and to better inform maternal-infant immunization practices, Front. Immunol., 6 (2015), 64. |
| [27] | WHO, Global tuberculosis report 2017. |
| [28] | European Centre for Disease Prevention and Control/WHO Regional Office for Europe, Tuberculosis surveillance and monitoring in Europe 2017. |
| [29] | K. Lönnroth, G. B. Migliori, I. Abubakar, et al., Towards tuberculosis elimination: an action framework for low-incidence countries, Eur. Respir. J., 45 (2015), 928–952. |
| [30] | RKI, Berichte zur Epidemiologie der Tuberkulose in Deutschland für 2016 (Reports on the epidemiology of tuberculosis in Germany for 2016). |
| [31] | B. Hauer and N. Perumal, Tuberkulose bleibt eine Herausforderung auch für Deutschland (Tuberculosis remains a challenge for Germany as well) (in german), Epid. Bull., 11/12 (2018), 109–111. |
| [32] | N. T. Mutters, F. Günther, A. Sander, et al., Influx of multidrug-resistant organisms by country-to-country transfer of patients, BMC Infect. Diseases, 15 (2015), 466–472. |
| [33] | A. Roy, M. Eisenhut, R. J. Harris, et al., Effect of BCG vaccination against Mycobacterium tuberculosis infection in children: systematic review and meta-analysis, BMJ, 349 (2014), g4643. |
| [34] | C. A. Thaissa and S. H. E. Kaufmann, Toward novel vaccines against tuberculosis: Current hopes and obstacles, Yale J. Biol. Med., 83 (2010), 209–215. |
| [35] | B. E. Kwon, J. H. Ahn, S. Min, et al., Development of new preventive and therapeutic vaccines for tuberculosis, Immune. Netw., 18 (2018), e17. |
| [36] | C. S. Merle, S. S. Cunha and L. C. Rodrigues, BCG vaccination and leprosy protection: review of current evidence and status of BCG in leprosy control, Expert Rev. Vaccines, 9 (2010), 209–222. |
| [37] | S. Chen, N. Zhang, J. Shao, et al., Maintenance versus non-maintenance intravesical Bacillus Calmette-Guérin instillation for non-muscle invasive bladder cancer: A systematic review and meta-analysis of randomized clinical trials, Int. J. Surg., 52 (2018), 248–257. |
| [38] | V. Nankabirwa, J. K. Tumwine, P. M. Mugaba, et al., Child survival and BCG vaccination: a community based prospective cohort study in Uganda, BMC Public Health., 15 (2015), 175–185. |
| [39] | M. G. Netea and R. van Crevel, BCG-induced protection: Effects on innate immune memory, Semin. Immunol., 26 (2014), 512–517. |
| [40] | B. Freyne, A. Marchant, N. Curtis, et al., BCG-associated heterologous immunity, a historical perspective: experimental models and immunological mechanisms, Trans. R. Soc. Trop. Med. Hyg., 109 (2015), 46–51. |
| [41] | P. Aaby, A. Roth, H. Ravn, et al., Randomized trial of BCG vaccination at birth to low-birth-weight children: beneficial nonspecific effects in the neonatal period?, J. Infect. Dis., 204 (2011), 245–252. |
| [42] | R. Kandasamy, M. Voysey, F. McQuaid, et al., Non-specific immunological effects of selected routine childhood immunisations: systematic review, BMJ, 355 (2016), i5225. |
| [43] | R. Ragonnet, J. Trauer, J. Denholm, et al., Vaccination programs for endemic infections: Modelling real versus apparent impacts of vaccine and infection characteristics, Nature Sci, Rep,, 5 (2015), 15468. |
| [44] | F. Shann, H. Nohynek, J. A. Scott, et al., Randomized trials to study the nonspecific effects of vaccines in children in low-income countries, Pediatr. Infect. Dis. J., 29 (2010), 457–461. |
| [45] | S. Sorup, M. Villumsen, H. Ravn, et al., Smallpox vaccination and all-cause infectious disease hospitalization: a Danish register-based cohort study, Int. J. Epidemiol., 40 (2011), 955-963. |
| [46] | S. Sorup, C. S. Benn, A. Poulsen, et al., Live vaccine against measles, mumps, and rubella and the risk of hospital admissions for nontargeted infections, JAMA, 311 (2014), 826-835. |
| [47] | WHO, Systematic review of the non-specific immunological effects of selected routine immunizations, Oxford University, 2015. |
| [48] | L. C. J. de Bree, V. A. C. M. Koeken, L. A. B. Joosten, et al., Non-specific effects of vaccines: Current evidence and potential implications, Semin. Immunol., 39 (2018), 35–43. |
| [49] | D. Uthayakumar, S. Paris, L. Chapat, et al., Non-specific Effects of Vaccines Illustrated Through the BCG Example: From Observations to Demonstrations, Front. Immunol., 9 (2018), Article 2869. |
| [50] | C. S. Benn, A. B. Fisker, A. Rieckmann, et al., How to evaluate potential non-specific effects of vaccines: the quest for randomized trials or time for triangulation?, Expert Rev. Vaccines, 17 (2018), 411–420. |
| [51] | M. J. de Castro, J. Pardo-Seco, F. Martinón-Torres, et al., Nonspecific (heterologous) protection of neonatal BCG vaccination against hospitalization due to respiratory infection and sepsis, Clin. Infect. Dis., 60 (2015), 1611–1619. |
| [52] | L. Sanders, S. Maiwald and H. Brunner, Epidemiologische Studie zu spezifischen und unspezifischen Wirkungen der BCG-Impfung in Deutschland mit Kosten-Nutzen-Bewertung (Epidemiological study on specific and non-specific effects of BCG vaccination in Germany with cost-benefit assessment) (in german), 62. Annual Conference of the German Society for Medical Informatics, Biometry and Epidemiology, Oldenburg 2017. |
| [53] | J. Leentjens, M. Kox, R. Stokman, et al., BCG vaccination enhances the immunogenicity of subsequent Influenza vaccination in healthy volunteers: a randomized, placebo-controlled pilot study, J. Infect. Dis., 212 (2015), 1930–1938. |
| [54] | K. L. Flanagan, S. L. Klein, N. E. Skakkebaek, et al., Sex differences in the vaccine-specific and non-targeted effects of vaccines, Vaccine, 29 (2011), 349–354. |
| [55] | L. G. Stensballe, S. Sørup, P. Aaby, et al., BCG vaccination at birth and early childhood hospitalisation: a randomised clinical multicentre trial, Arch. Dis. Child., 102 (2017), 224–231. |
| [56] | L. G. Stensballe, H. Ravn, N. Birk, et al., BCG vaccination at birth and rate of hospitalization for Infection until 15 months of age in Danish children: A randomized clinical multicenter trial, J. Pediatric Infect. Dis. Soc., 2018. |
| [57] | A. Rieckmann, M. Villumsen, M. L. Jensen, et al., The effect of smallpox and Bacillus Calmette-Guérin vaccination on the risk of Human Immunodeficiency Virus-1 infection in Guinea-Bissau and Denmark, Open Forum Infect. Dis., 4 (2017), ofx 130. |
| [58] | S. Prentice, E. L. Webb, H. M. Dockrell, et al., Investigating the non-specific effects of BCG vaccination on the innate immune system in Ugandan neonates: study protocol for a randomised controlled trial, Trials, 16 (2015), 149–161. |
| [59] | C. S. Benn, A. B. Fisker, H. C. Whittle, et al., Revaccination with live attenuated vaccines confer additional beneficial nonspecific effects on overall survival: A review, EBioMedicine, 10 (2016), 312–317. |
| [60] | N. M. Birk, T. N. Nissen, J. Kjægaard, et al., Effects of Bacillus Calmette-Guérin (BCG) vaccination at birth on T and B lymphocyte subsets: Results from a clinical randomized trial, Sci. Rep., 7 (2017), 12398. |
| [61] | S. Saeed, J. Quintin, H. H. Kerstens, et al., Epigenetic programming of monocyte-to-macrophage differentiation and trained innate immunity, Science, 345 (2014), 1251086. |
| [62] | J. Kleinnijenhuis, J. Quntin, F. Preijers, et al., Bacille Calmette-Guérin induces NOD2-dependent nonspecific protection from reinfection via epigenetic reprogramming of monocytes, Proc. Natl. Acad. Sci. USA, 109 (2012), 17537–17542. |
| [63] | Y. L. Lee, J. Y. Lee, Y. H. Jang, et al., Non-specific effects of vaccines: immediate protection against Respiratory Syncytial Virus infection by Line Attenuated Influenza Vaccine, Front. Microbiol., 9 (2018), Article 83. |
| [64] | R. J. W. Arts, S. J. C. F. M. Moorlag, B. Novakovic, et al., BCG Vaccination Protects against Experimental Viral Infection in Humans through the Induction of Cytokines Associated with Trained Immunity, Cell Host & Microbe., 23 (2018), 89–100.e5. |
| [65] | B. Freyne, S. Donath, S. Germano, et al., Neonatal BCG vaccination influences cytokine responses to Toll-like Receptor ligands and heterologous antigens, J. Infect. Dis., 217 (2018), 1798–1808. |
| [66] | G. A. Weinberg and P. G. Szilagy, Vaccine epidemiology: Efficacy, effectiveness, and the translational research road map, J. Infect. Dis., 201 (2010), 1607–1610. |
| [67] | M. L. McHugh, The odds ratio: calculation, usage, and interpretation, Biochemica Medica, 19 (2009), 120–126. |
| [68] | S. Gao, Z. Teng, J. Nieto, et al., Analysis of an SIR Epidemic Model with Pulse Vaccination and Distributed Time Delay, J. Biomed. Biotechnol., 2007 (2007), 64870. |
| [69] | C. Castillo-Chavez and B. Song, Dynamical models of tuberculosis and their applications, Math. Biosci. Eng., 1 (2004), 361–404. |
| [70] | C. Ozcaglar, A Shabbear, S. L. Vandenberg, et al., Epidemiological models of Mycobacterium tuberculosis complex infections, Math. Biosci., 236 (2012), 77-9-6. |
| [71] | V. P. Driessche and J. Watmough, Reproduction numbers and subthreshold endemic equilibria for compartmental models of disease transmission, Math. Biosci., 180 (2002), 29–48. |
| [72] | WHO, Surveillance Report, 2012–2017, available from: https://ecdc.europa.eu/sites/portal/files/media/en/publications/Publications/ecdc-tuberculosis-surveillance-monitoring-Europe-2017.pdf |
| [73] | WHO, Vaccine-preventable diseases: monitoring system 2018 global summary, available from: http://apps.who.int/immunization_monitoring/globalsummary/timeseries/ tswucoveragebcg.html |
| [74] | K. Styblo and J. Meijer, Impact of BCG vaccination programmes in children and young adults on the tuberculosis problem, Tubercle, 57 (1976), 17–43. |
| [75] | J. Kleinnijenhuis, J. Quintin, F. Preijers, et al., Long-Lasting Effects of BCG Vaccination on Both Heterologous Th1/Th17 Responses and Innate Trained Immunity, J. Innate. Immun., 6 (2014), 152–158. |
| [76] | Health Data, South Sudan, available from: http://www.healthdata.org/south-sudan |
| [77] | Health Data, Guinea-Bissau, available from: http://www.healthdata.org/guinea-bissau |
| [78] | Index Mundi, Country Comparison: Guinea-Bissau vs. South Sudan, available from: http: //www.indexmundi.com/factbook/compare/guinea-bissau.south-sudan |
| [79] | WHO, Vaccine-preventable diseases: monitoring system 2018 global summary, Guinea Bissau, available from: http://apps.who.int/immunization_monitoring/globalsummary/estimates?c=GNB |
| [80] | WHO, Vaccine-preventable diseases: monitoring system 2018 global summary, South Sudan, available from: http://apps.who.int/immunization_monitoring/globalsummary/coverages?c=SSD |
| [81] | UN, available from: http://www.childmortality.org/files_v20/download/IGME% 20Report%202015_9_3%20LR%20Web.pdf (Inter-agency Group for Child Mortality Estimation, 2010–2015) |
| [82] | UNICEF, available from: http://data.unicef.org/resources/state-worlds-children-2016-statistical-tables/ (2007–2016) |
| [83] | P. Mangtani, P. Nguipdop-Djomo, R. H. Keogh, et al., The duration of protection of school-aged BCG vaccination in England: a population-based case–control study, Int. J. Epidemiol., 47 (2017), 193–201. |
| [84] | D. P. Gao and N. J. Huang, A note on global stability for a tuberculosis model, Appl. Math. Lett., 73 (2017), 163–168. |
| [85] | D. P. Gao and N. J. Huang, Optimal control analysis of a tuberculosis model, Appl. Math. Modell., 58 (2018), 47–64. |
| [86] | H.-F. Huo, S.-J. Dang and Y.-N. Li, Stability of a two-strain tuberculosis model with general contact rate, Abstr. Appl. Anal., 2010 (2010), Article ID 293747. |
| [87] | H.-F. Huo and L.-X. Feng, Global Stability of an Epidemic Model with Incomplete Treatment and Vaccination, Discrete Dyn. Nat. Soc., 2012, Article ID 530267. |
| [88] | J. Liu and T. Zhang, Global stability for a tuberculosis model, Math. Comput. Modell., 54 (2011), 836–845. |
| [89] | C. Vargas-De-Leon, On the global stability of infectious diseases models with relapse, Abstraction & Application, 9 (2013), 50–61. |
| [90] | W. Wojtak, C. J. Silva and D. F. M. Torres, Uniform asymptotic stability of a fractional tuberculosis model, Math. Modell. Natur. Phenom., 13 (2018). |
| [91] | R. Chinnathambi, F. A. Rihan and H. J. Alsakaji, A fractional-order model with time delay for tuberculosis with endogenous reactivation and exogenous reinfections, Math. Meth. Appl. Sci., May 2019. |
| [92] | S. Bowong and A. M. A. Alaoui, Optimal intervention strategies for tuberculosis, Commun. Nonlin. Sci. Numer. Simul., 18 (2013), 1441–1453. |
| [93] | L. J. S. Allen, An Introduction to Stochastic Epidemic Models, in: F. Brauer, P. van den Driessche, J. Wu (eds.), Mathematical Epidemiology. Lecture Notes in Mathematics 1945, Springer, Berlin, Heidelberg, pp 81–130. |
| [94] | J. L. Dimi and T. Mbaya, Dynamics analysis of stochastic tuberculosis model transmission with immune response, AIMS Math., 3 (2018), 391–408. |
| [95] | A. El Myr, A. Assadouq, L. Omari, et al., A Stochastic SIR Epidemic System with a Nonlinear Relapse, Discrete Dyn. Nat. Soc., 2018 (2018), Article ID 5493270. |
| [96] | T. Feng and Y. Qiu, Global analysis of a stochastic TB model with vaccination and treatment, Discrete & Cont. Dyn-B, 24 (2019), 2923–2939. |
| [97] | Q. Liu, D. Jiang, T. Hayat, et al., Dynamics of a stochastic tuberculosis model with antibiotic resistance, Chaos Solitons & Fract., 109 (2018), 223–230. |
| [98] | Q. Liu and D. Jiang, The dynamics of a stochastic vaccinated tuberculosis model with treatment, Physica A, 527 (2019), 121274. |
| [99] | M. Mbokoma and S. C. O. Noutchie, Mathematical analysis of a stochastic tuberculosis model, J. Anal. Appl., 15 (2017), 21–50. |
| [100] | B. Song, C. Castillo-Chavez andJ. P. Aparicio, Tuberculosis models withfast and slowdynamics: The role of close and casual contacts, Math. Biosci., 180 (2002), 187–205. |
| [101] | D. Bichara and A. Iggidr, Multi-patch and multi-group epidemic models: a new framework, J. Math. Biol., 77 (2018), 107–134. |
| [102] | E. Pienaar, A. M. Fluitt, S. E. Whitney, et al., A model of tuberculosis transmission and intervention strategies in an urban residential area, Comput. Biol. Chem., 34 (2010), 86–96. |
| [103] | X. Hu, Threshold dynamics for a Tuberculosis model with seasonality, Math. Biosci. Eng., 9 (2012), 111–122. |
| [104] | L. Liu, X.-Q. Zhao and Y. Zhou, A Tuberculosis Model with Seasonality, Bull. Math. Biol., 72 (2010), 931–952. |
| [105] | H.Xiang, M.-X.ZouandH.-F.Huo, ModelingtheEffectsofHealthEducationandEarlyTherapy on Tuberculosis Transmission Dynamics, Int. J. Nonlin. Sci. Numer. Simul., March 2019. |
| [106] | N. Blaser, C. Zahnda, S. Hermans, et al., Tuberculosis in Cape Town: An age-structured transmission model, Epidemics, 14 (2016), 54–61. |
| [107] | R. Xu, Global dynamics of an epidemiological model with age of infection and disease relapse, J. Biol. Dynam., 12 (2017), 118–145. |
| [108] | L. Liu, X. Ren and Z. Jin, Threshold dynamical analysis on a class of age-structured tuberculosis model with immigration of population, Adv. Diff. Eqs., 2017 (2017), 258. |
| [109] | S. Khajanchi, D. K. Das and T. K. Kar, Dynamics of tuberculosis transmission with exogenous reinfections and endogenous reactivation, Physica A, 497 (2018), 52–71. |
| [110] | B. K. Mishra and J. Srivastava, Mathematical model on pulmonary and multidrug-resistant tuberculosis patients with vaccination, J. Egypt. Math. Soc., 22 (2014), 311–316. |
| [111] | H.-F. Huo and M.-X. Zou, Modelling effects of treatment at home on tuberculosis transmission dynamics, Appl. Math. Modell., 40 (2016), 9474–9484. |
| [112] | M. J. Keeling and K. T. D. Eames, Networks and epidemic models, J. R. Soc. Interface, 2 (2005), 295–307. |
| [113] | M. E. J. Newman, Spread of epidemic disease on networks, Phys. Rev. E, 66 (2002), 016128. |
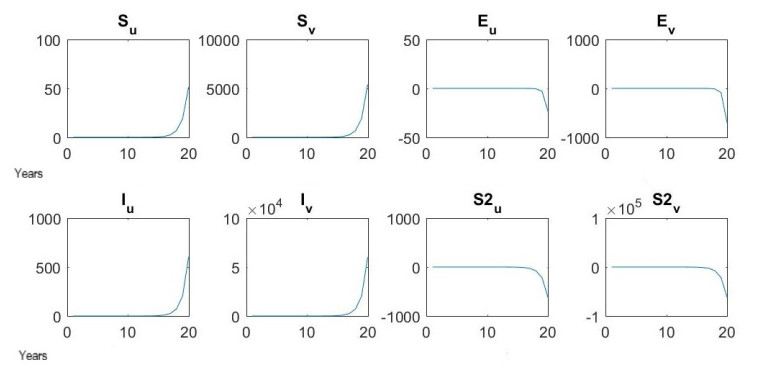
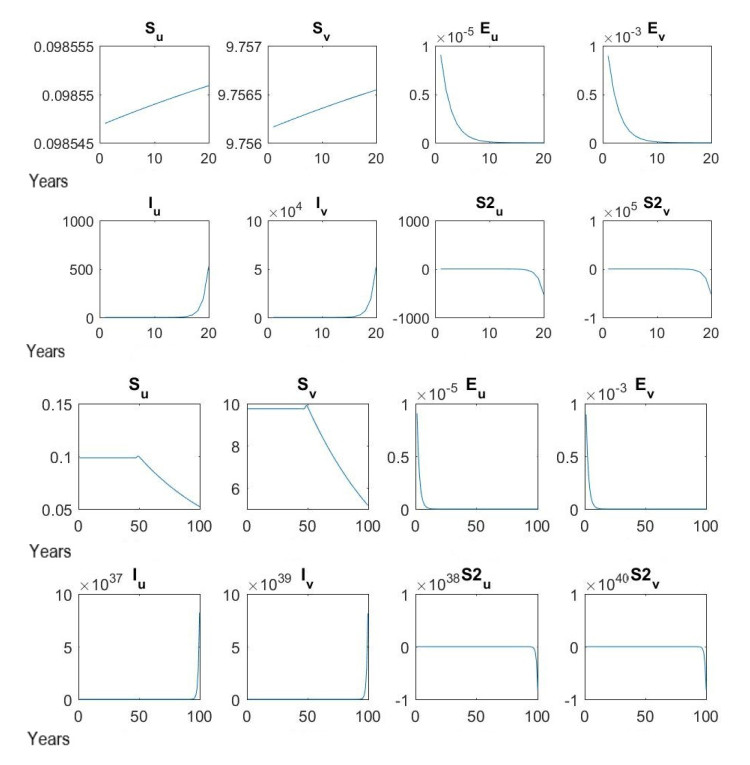
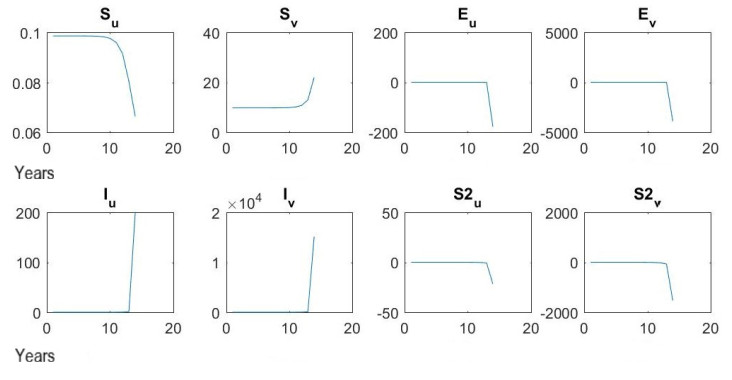
Figures(7) / Tables(16)

Sarah Treibert, Helmut Brunner, Matthias Ehrhardt. Compartment models for vaccine effectiveness and non-specific effects for Tuberculosis[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 7250-7298. doi: 10.3934/mbe.2019364







 DownLoad:
DownLoad: