Citation: Chunguang Dong, Zhuojie Zhang, Xiaoxia Zhen, Mu Chen. Free vibration of summation resonance of suspended-cable-stayed beam[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 7230-7249. doi: 10.3934/mbe.2019363

| [1] | K. Takahashi and Y. Konishi, Non-linear vibrations of cables in three dimensions, Part II: out-of-plane vibrations under in-plane sinusoidally time-varying load, J. Sound Vib., 118 (1987), 85–97. |
| [2] | J. L. Lilien and A. P. Da Costa, Vibration amplitudes caused by parametric excitation of cable stayed structures, J. Sound Vib., 174 (1994), 69–90. |
| [3] | H. N. Arafat and A. H. Nayfeh, Non-linear responses of suspended cables to primary resonance excitations, J. Sound Vib., 266 (2003), 325–354. |
| [4] | H. Wang, T. Y. Tao, R. Zhou, et al., Parameter sensitivity study on flutter stability of a long-span triple-tower suspension bridge, J. Wind Eng. Ind. Aerod., 128 (2014), 12–21. |
| [5] | T. Grigorjeva and Z. Kamaitis, Numerical analysis of the effects of the bending stiffness of the cable and the mass of structural members on free vibrations of suspension bridges, J. Civ. Eng. Manag., 21 (2015), 948–957. |
| [6] | C. G. Koh and Y. Rong, Dynamic analysis of large displacement cable motion with experimental verification, J. Sound Vib., 272 (2004), 183–206. |
| [7] | M. H. EI Ouni and N. B. Kahla, Nonlinear dynamic analysis of a cable under first and second order parametric excitations, J. Civ. Eng. Manag., 18 (2012), 557–567. |
| [8] | D. Xue, Y. Liu, J. He, et al., Experimental study and numerical analysis of a composite truss joint, J. Constr Steel Res., 67 (2011), 957–964. |
| [9] | K. Takahashi, Dynamic stability of cables subjected to an axial periodic load, J. Sound Vib., 144 (1991), 323–330. |
| [10] | A. P. Da Costa and J. A. C. Martins, Oscillations of bridge stay cables induced by periodic motions of deck and/or towers, J. Eng. Mech., 122 (1996), 613–622. |
| [11] | H. Chen, D. Zuo, Z. Zhang, et al., Bifurcations and chaotic dynamics in suspended cables under simultaneous parametric and external excitations, Nonlinear Dyn., 62 (2010), 623–646. |
| [12] | D. Zulli and A. Luongo, Nonlinear energy sink to control vibrations of an internally nonresonant elastic string, Meccanica, 50 (2014), 781–794. |
| [13] | B. N. Sun and Z. G. Wang, Cable oscillation induced by parametric excitation in cable-stayed bridges, J. Zhejiang Univ-sc A, 4 (2003), 13–20. |
| [14] | Y. Fujino and P. Warnitchai, An experimental and analytical study of auto parametric resonance in a 3-DOF model of cable-stayed-beam, Nonlinear Dyn., 4 (1993), 111–138. |
| [15] | Y. Xia and Y. Fujino, Auto-parametric vibration of a cable-stayed-beam structure under random excitation, J. Eng. Mech., 132 (2006), 279–286. |
| [16] | Y. Xia, Q. X. Wu, Y. L. Xu, et al., Verification of a cable element for cable parametric vibration of one-cable-beam system subject to harmonic excitation and random excitation, Adv. Struct. Eng., 14 (2011), 589–595. |
| [17] | V. Gattulli and M. Lepidi, Nonlinear interactions in the planar dynamics of cable-stayed beam, Int J. Solids Struct., 40 (2003), 4729–4748. |
| [18] | V. Gattulli and M. Morandini, A parametric analytical model for non‐linear dynamics in cable-stayed beam, Earthq. Eng. Struct. D., 31 (2002), 1281–1300. |
| [19] | V. Gattulli and M. Lepidi, One-to-two global-local interaction in a cable-stayed beam observed through analytical, finite element and experimental models, Int. J. Nonlin Mech., 40 ( 2005), 571–588. |
| [20] | Z. Wang, C. Sun, Y. Zhao, et. al., Modeling and nonlinear modal characteristics of the cable-stayed beam, Eur. J. Mech- A-Solid, 47 (2014), 58–69. |
| [21] | M. H. Wei, K. Lin, L. Jin, et. al., Nonlinear dynamics of a cable-stayed beam driven by sub-harmonic and principal parametric resonance, Int. J. Mech. Sci., 110 (2016), 78–93. |
| [22] | Z. W. Wang and T. J. Li, Nonlinear dynamic analysis of parametrically excited space cable-beam structures due to thermal loads, Eng. Struct., 83 (2015), 50–61. |
| [23] | H. J. Kang, H. P. Zhu, Y. Y. Zhao, et al., In-plane non-linear dynamics of the stay cables, Nonlinear Dyn., 73 (20136), 54–59. |
| [24] | Y. Y. Zhao and H. J. Kang, In-plane free vibration analysis of cable-arch structure, J. Sound Vib., 312 (2008), 363–379. |
| [25] | H. J. Kang, Y. Y. Zhao and H. P. Zhu, Out-of-plane free vibration analysis of a cable-arch structure, J. Sound Vib., 332 (2013), 907–921. |
| [26] | A. H. Nayfeh and D. T. Mook, Nonlinear Oscillations, John Wiley & Sons, (1979), 71–79. |
| [27] | J. R. Zhao, Y. G. Tang and L. Q. Liu, Study on the sum type combination resonance responses of a classic Spar platform, Ocean Eng., 27 (2009), 23–30. |
| [28] | L. Q. Chen, J. W. Zu and J. Wu, Steady-state response of the parametrically excited axially moving string constituted by the Boltzmann superposition principle, Acta Mech., 162 (2003), 143–155. |
| [29] | Q. X. Wu, L. Liu and B. C. Chen, Theoretical equations of in-plane natural vibration for cables considering bending stiffness, Eng. Mech., 27 (2010), 9–27. |
| [30] | M. Irvine, Cable Structures, The MIT Series in Structural Mechanics, (1981). |
| [31] | Japan Society of Civil Engineers, Basic and application of cable and space structures, Maruzen, Japan: JSCE, (1999). |
| [32] | G. H. Li, H. F. Xiang and Z. Y. Shen, Stability and vibration of bridge structures, China Railway Publishing House, (2010), 132–134. |
| [33] | N. J. Gimsing and C. T. Georgakis, Cable supported bridges: concept and design. 3th ed, John Wiley & Sons, (2011), 220–225. |
| [34] | A. H. Nayfeh and D. T. Mook, Nonlinear Oscillation, John Wiley & Sons, (1979). |
| [35] | M. Al-Qassab and S. Nair, Wavelet-Galerkin method for the free vibrations of an elastic cable carrying an attached mass, J. Sound Vib., 270 (2004), 191–206. |
| [36] | S. H. Zhou, G. Q. Song, Z. H. Ren, et al., Nonlinear analysis of a parametrically excited beam with intermediate support by using Multi-dimensional incremental harmonic balance method, Chaos, Solitons & Fractals, 93 (2016), 207–222. |
| [37] | P. J. Li, R. H. Wang and N. J. Ma, Analytic algorithm of transverse vibration frequency of cables with intermediate flexible supports, J. South China Univ T., 40 (2012), 7–18. |
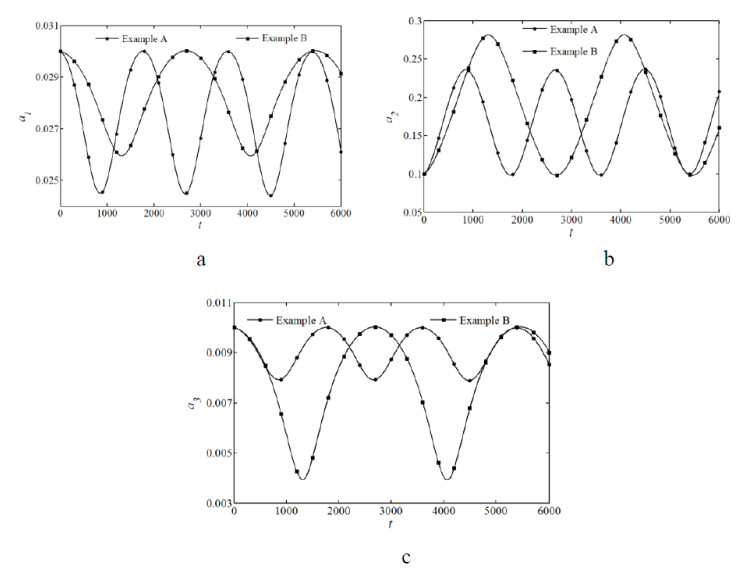
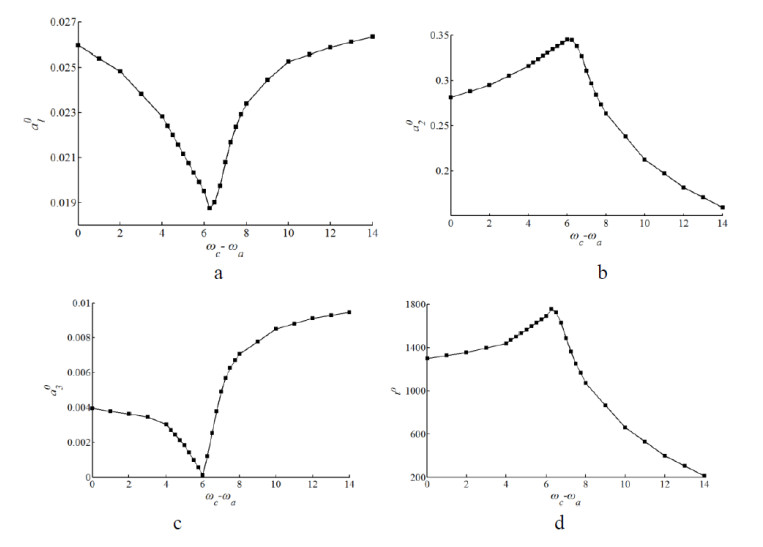
Figures(10) / Tables(3)

Chunguang Dong, Zhuojie Zhang, Xiaoxia Zhen, Mu Chen. Free vibration of summation resonance of suspended-cable-stayed beam[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 7230-7249. doi: 10.3934/mbe.2019363







 DownLoad:
DownLoad: