Citation: Luis Almeida, Michel Duprez, Yannick Privat, Nicolas Vauchelet. Mosquito population control strategies for fighting against arboviruses[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 6274-6297. doi: 10.3934/mbe.2019313

| [1] | A. A. Hoffmann, B. L. Montgomery, J. Popovici, et al., Successful establishment of Wolbachia in Aedes populations to suppress dengue transmission, Nature, 476 (2011), 454–457. |
| [2] | B. Stoll, H. Bossin, H. Petit, et al., Suppression of an isolated population of the mosquito vector Aedes polynesiensis on the atoll of Tetiaroa, French Polynesia, by sustained release of Wolbachia-incompatible male mosquitoes, Conference: ICE - XXV International Congress of Entomology, At Orlando, Florida, USA. |
| [3] | D. D. Thomas, C. A. Donnelly, R. J. Wood, et al., Insect population control using a dominant, repressible, lethal genetic system. Science, 287 (2000), 2474–2476. |
| [4] | J. Heinrich and M. Scott, A repressible female-specific lethal genetic system for making transgenic insect strains suitable for a sterile-release program. Proc. Natl. Acad. Sci. USA, 97 (2000), 8229–8232. |
| [5] | G. Fu, R. S. Lees, D. Nimmo, et al., Female-specific flightless phenotype for mosquito control Proc. Natl. Acad. Sci., 107 (2010), 4550–4554. |
| [6] | F. Gould, Y. Huang, M. Legros, et al., A Killer–Rescue system for self-limitinggene drive of anti-pathogen constructs, Proc. R. Soc. B, 275 (2008), 2823–2829. |
| [7] | J. M. Marshall, G. W. Pittman, A. B. Buchman, et al., Semele: a killer-male, rescue-female system for suppression and replacement of insect disease vector populations, Genetics, 187 (2011), 535–551. |
| [8] | C. M. Ward, J. T. Su, Y. Huang, et al., Medea selfish genetic elements as tools for altering traits of wild populations: a theoretical analysis. Evolution, 65 (2011), 1149–1162. |
| [9] | M. A. Robert, K. Okamoto, F. Gould, et al., A reduce and replace strategy for suppressing vector-borne diseases: insights from a deterministic model, PLoS ONE, 8 (2012), e73233. |
| [10] | H.J. Barclay and M. Mackuer, The sterile insect release method for pest control: a density dependent model, Environ. Entomol., 9 (1980), 810–817. |
| [11] | V.A.Dyck, J.HendrichsandA.S.Robinson, TheSterileInsectTechnique, PrinciplesandPractice in Area-Wide Integrated Pest Management, Springer, Dordrecht, 2006 |
| [12] | R. Anguelov, Y. Dumont and J. Lubuma, Mathematical modeling of sterile insect technology for control of anopheles mosquito, Comput. Math. Appl., 64 (2012), 374–389. |
| [13] | C. Dufourd and Y. Dumont, Impact of environmental factors on mosquito dispersal in th e prospect of sterile insect technique control, Comput. Math. Appl., 66 (2013), 1695–1715. |
| [14] | Y. Dumont and J. M. Tchuenche, Mathematical studies on the sterile insect technique for the chikungunya disease and Aedes albopictus, J. Math. Biol., 65 (2012), 809–855. |
| [15] | L. Cai, S. Ai and J. Li, Dynamics of mosquitoes populations with different strategies for releasing sterile mosquitoes, SIAM J. Appl. Math., 74 (2014), 1786–1809. |
| [16] | J. Li and Z. Yuan, Modelling releases of sterile mosquitoes with different strategies, J. Biol. Dyn., 9 (2015), 1–14. |
| [17] | G. Sallet and M. A. H. B. da Silva, Monotone dynamical systems and some models of Wolbachia in aedes aegypti populations, ARIMA, 20 (2015), 145–176. |
| [18] | M. Huang, X. Song and J. Li, Modelling and analysis of impulsive releases of sterile mosquitoes, J. Biol. Dyn., 11 (2017), 147–171. |
| [19] | H. Bossin, Y. Dumont and M. Strugarek, Using sterilizing males to reduce or eliminate Aedes populations: insights from a mathematical model, Appl. Math. Model., 68 (2019), 443–470. |
| [20] | P.-A. Bliman, D. Cardona-Salgado, Y. Dumont, et al., Implementation of Control Strategies for Sterile Insect Techniques, arXiv:1812.01277. |
| [21] | K. Bourtzis, Wolbachia-based technologies for insect pest population control, In: Aksoy S. (eds) Transgenesis and the Management of Vector-Borne Disease. Advances in Experimental Medicine and Biology, 627 (2008), Springer, New York, NY. |
| [22] | S. P. Sinkins, Wolbachia and cytoplasmic incompatibility in mosquitoes, Insect Biochem. Mol. Biol., 34 (2004), 723–729. |
| [23] | J. H. Werren, L. Baldo and M. E. Clark, Wolbachia: master manipulators of invertebrate biology, Nature Rev. Microbiol., 8 (2008), 741–751. |
| [24] | T. Walker, P. H. Johnson, L. A. Moreira, et al., The wMel Wolbachia strain blocks dengue and invades caged Aedes aegypti populations, Nature, 476 (2011), 450–453. |
| [25] | J. Z. Farkas and P. Hinow, Structured and unstructured continuous models for wolbachia infections, B. Math. Biol., 72 (2010), 2067–2088. |
| [26] | A. Fenton, K. N. Johnson, J. C. Brownlie, et al., Solving the Wolbachia paradox: modeling the tripartite interaction between host, Wolbachia, and a natural enemy, The American Naturalist, 178 (2011), 333–342. |
| [27] | G. Schraiber, A. N. Kaczmarczyk, R. Kwok, et al., Constraints on the use of lifespan-shortening wolbachia to control dengue fever, J. Theor. Bio., 297 (2012), 26–32. |
| [28] | H. Hughes and N. F. Britton, Modeling the Use of Wolbachia to Control Dengue Fever Transmission, Bull. Math. Biol., 75 (2013), 796–818. |
| [29] | P.-A. Bliman, M. S. Aronna, F. C. Coelho, et al., Ensuring successful introduction of Wolbachia in natural populations of Aedes aegypti by means of feedback control, J. Math. Biol., 76 (2018), 1269–1300. |
| [30] | M. Strugarek, Modélisation mathématique de dynamiques de populations, applications à la lutte anti-vectorielle contre Aedes spp. (Diptera:Culicidae), Ph.D thesis, Sorbonne Université, 2018. |
| [31] | R. C. A. Thome, H. M. Yang and L. Esteva, Optimal control of Aedes aegypti mosquitoes by the sterile insect technique and insecticide, Math. Biosci., 223 (2010), 12–23. |
| [32] | D. E. Campo-Duarte, O. Vasilieva, D. Cardona-Salgado, et al., Optimal control approach for establishing wMelPop Wolbachia infection among wild Aedes aegypti populations, J. Math. Biol., 76 (2018), 1907–1950. |
| [33] | P.-A. Bliman, Feedback Control Principles for Biological Control of Dengue Vectors, preprint. |
| [34] | H. Yang, M. Macoris, K. Galvani, et al., Assessing the effects of temperature on the population of Aedes aegypti, the vector of dengue, Epidemiol. Infect., 137 (2009), 1188–1202. |
| [35] | A. A. Hoffmann and M. Turelli, Cytoplasmic incompatibility in insects, pp 42-80 in S. L. O'Neil, J. H. Werren, and A. A. Hoffmann, eds, Influential passengers: inherited microorganisms and arthropod reproduction. Oxford University Press, Oxford. |
| [36] | M. Strugarek and N. Vauchelet, Reduction to a single closed equation for 2 by 2 reaction-diffusion systems of Lotka-Volterra type, SIAM J. Appl. Math., 76 (2016), 2068–2080. |
| [37] | L. Almeida, Y. Privat, M. Strugarek, et al., Optimal releases for population replacement strategies, application to Wolbachia, to appear in SIAM J. Math. Anal., preprint, Hal-01807624 (2018). |
| [38] | L. Beal, D. Hill, R. Martin, et al., GEKKO Optimization Suite, Processes, Multidisciplinary Digital Publishing Institute, 6 (2018), 106. |
| [39] | A. Wächter and L. T. Biegler. On the implementation of an interior-point filter line-search algorithm for large-scale nonlinear programming. Math. Program., 106 (2006), 25–57. |
| [40] | H. Dutra, L. dos Santos, E. Caragata, et al., From lab to field: the influence of urban landscapes on the invasive potential of Wolbachia in Brazilian Aedes aegypti mosquitoes, PLoS Neglect. Rrop. D., 9 (2015), e0003689. |
| [41] | L. Almeida, M. Duprez, Y. Privat, et al., Optimal release strategy for the sterile mosquitoes technique, work in progress. |
| [42] | E. B. Lee and L. Markus, Foundations of optimal control theory, SIAM Series Appl. Math., Wiley, 1967. |
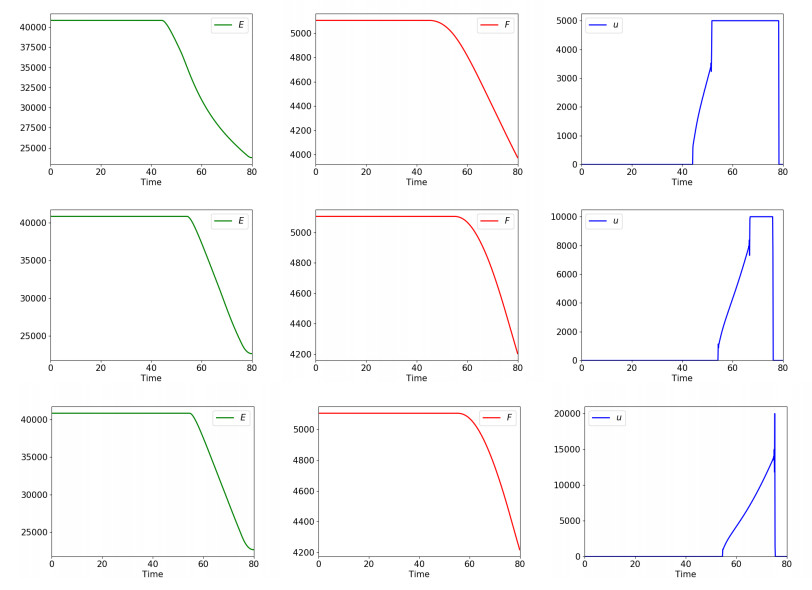
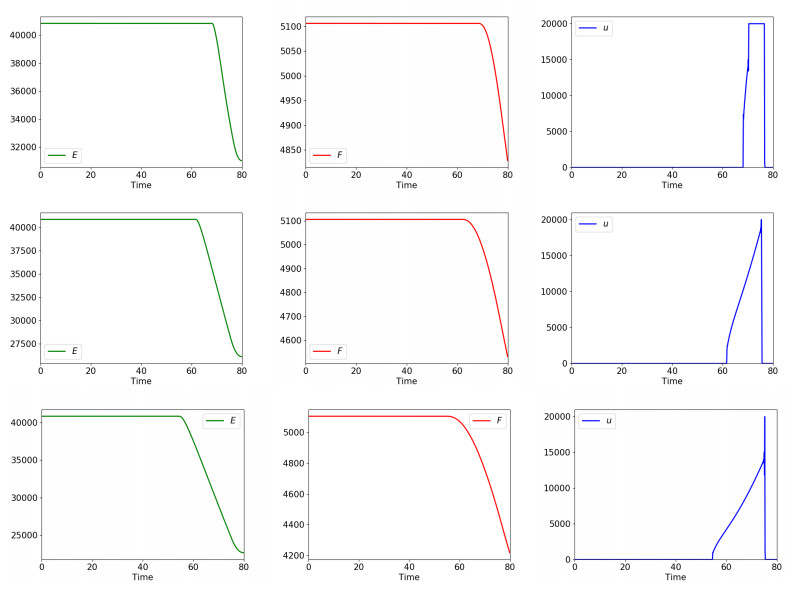
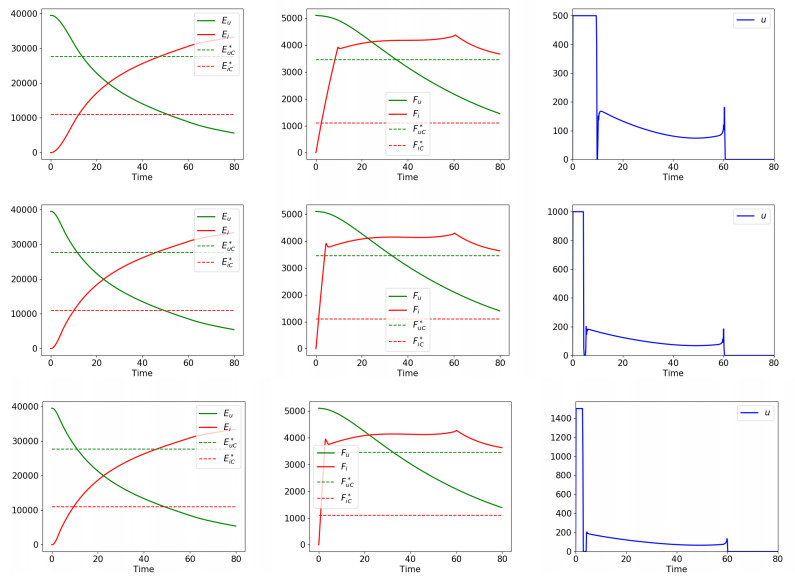
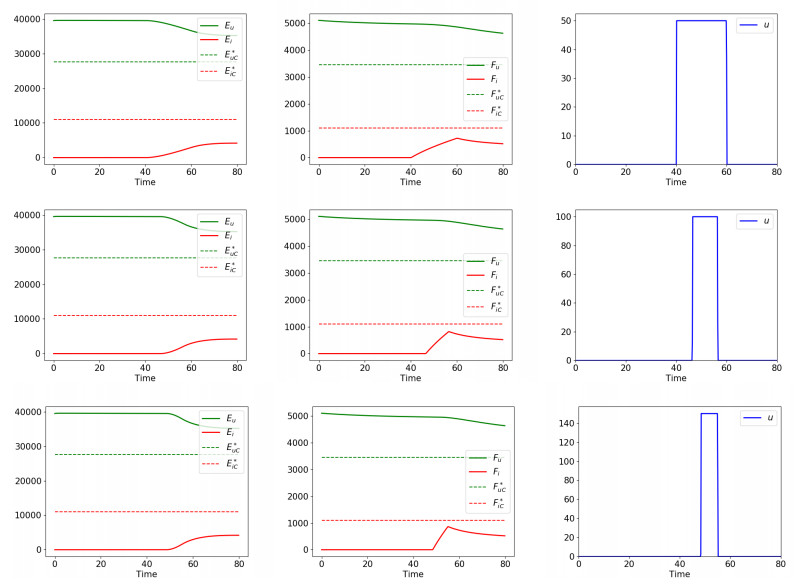
Figures(5) / Tables(2)

Luis Almeida, Michel Duprez, Yannick Privat, Nicolas Vauchelet. Mosquito population control strategies for fighting against arboviruses[J]. Mathematical Biosciences and Engineering, 2019, 16(6): 6274-6297. doi: 10.3934/mbe.2019313







 DownLoad:
DownLoad: