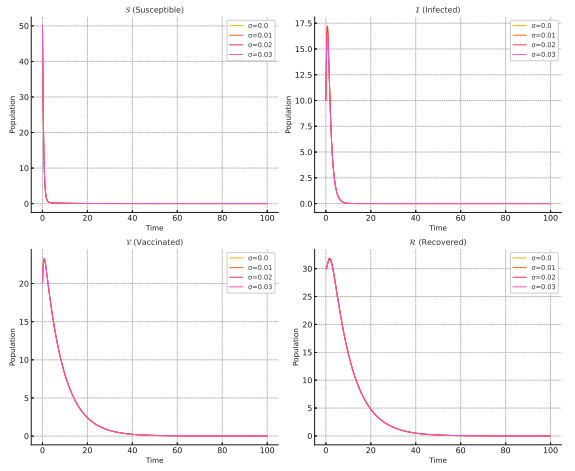
Effective disease control measures are essential for mitigating epidemic risks. This study introduces a novel stochastic susceptible-infected-vaccinated-recovered $ \mathscr{SIVR} $ epidemic model that incorporates white noise in vaccination dynamics. Unlike traditional deterministic models, our stochastic framework accounts for the inherent randomness in real-world disease transmission and the effectiveness of interventions. We rigorously establish the existence and uniqueness of global positive solutions using Lyapunov functions and derive conditions for disease extinction and persistence under stochastic perturbations. A key contribution is the introduction of a stochastic reproduction number $R^*_0$, which refines classical epidemic thresholds by integrating randomness. Through numerical simulations, we illustrate the impact of stochasticity on disease dynamics, demonstrating that noise can drive disease extinction even in scenarios where deterministic models predict persistence. This study provides a more realistic epidemiological framework for optimizing vaccination strategies under uncertainty, offering significant advances in epidemic modeling and public health policy.
Citation: Shah Hussain, Naveed Iqbal, Elissa Nadia Madi, Thoraya N. Alharthi, Ilyas Khan. Vaccination strategies in a stochastic $ \mathscr{SIVR} $ epidemic model[J]. AIMS Mathematics, 2025, 10(2): 4441-4456. doi: 10.3934/math.2025204
Effective disease control measures are essential for mitigating epidemic risks. This study introduces a novel stochastic susceptible-infected-vaccinated-recovered $ \mathscr{SIVR} $ epidemic model that incorporates white noise in vaccination dynamics. Unlike traditional deterministic models, our stochastic framework accounts for the inherent randomness in real-world disease transmission and the effectiveness of interventions. We rigorously establish the existence and uniqueness of global positive solutions using Lyapunov functions and derive conditions for disease extinction and persistence under stochastic perturbations. A key contribution is the introduction of a stochastic reproduction number $R^*_0$, which refines classical epidemic thresholds by integrating randomness. Through numerical simulations, we illustrate the impact of stochasticity on disease dynamics, demonstrating that noise can drive disease extinction even in scenarios where deterministic models predict persistence. This study provides a more realistic epidemiological framework for optimizing vaccination strategies under uncertainty, offering significant advances in epidemic modeling and public health policy.

| [1] | R. Anderson, R. May, Infectious diseases of humans dynamics and control, 1 Eds., Oxford University Press, 1991. https://doi.org/10.1093/oso/9780198545996.001.0001 |
| [2] |
H. Hethcote, The mathematics of infectious diseases, SIAM Rev., 42 (2000), 599–653. https://doi.org/10.1137/S0036144500371907 doi: 10.1137/S0036144500371907

|
| [3] | F. Brauer, C. C. Chavez, Mathematical models in population biology and epidemiology, 2 Eds., New York: Springer, 2012. http://dx.doi.org/10.1007/978-1-4614-1686-9 |
| [4] |
F. Rihan, J. Alsakran, Dynamics of a stochastic delay differential model for COVID-19 infection with asymptomatic infected and interacting people: Case study in the UAE, Results Phys., 28 (2021), 104658. https://doi.org/10.1016/j.rinp.2021.104658 doi: 10.1016/j.rinp.2021.104658
|
| [5] | M. Keeling, P. Rohani, Modeling infectious diseases in humans and animals, Princeton University Press, 2011. https://doi.org/10.2307/j.ctvcm4gk0 |
| [6] |
A. Alkhazzan, J. Wang, Y. Nie, H. Khan, J. Alzabut, A stochastic Susceptible-Vaccinees-Infected-Recovered epidemic model with three types of noises, Math. Method. Appl. Sci., 47 (2024), 8748–8770. https://doi.org/10.1002/mma.10042 doi: 10.1002/mma.10042
|
| [7] |
A. Alkhazzan, J. Wang, Y. Nie, H. Khan, J. Alzabut, An effective transport-related SVIR stochastic epidemic model with media coverage and Levy noise, Chaos Soliton. Fract., 175 (2023), 113953. https://doi.org/10.1016/j.chaos.2023.113953 doi: 10.1016/j.chaos.2023.113953
|
| [8] |
J. Jacquez, P. O'Neill, Reproduction numbers and thresholds in stochastic epidemic models I. Homogeneous populations, Math. Biosci., 107 (1991), 161–186. https://doi.org/10.1016/0025-5564(91)90003-2 doi: 10.1016/0025-5564(91)90003-2
|
| [9] |
H. Alsakjia, A. Rihan, A. Hashish, Dynamics of a stochastic epidemic model with vaccination and multiple time-delays for COVID-19 in the UAE, Complexity, 1 (2022), 4247800. https://doi.org/10.1155/2022/4247800 doi: 10.1155/2022/4247800
|
| [10] | A. Din, Y. Li, A. Omame, A stochastic stability analysis of an HBV-COVID-19 coinfection model in resource limitation settings, Wave. Random Complex, 2022. https://doi.org/10.1080/17455030.2022.2147598 |
| [11] |
O. Diekmann, J. A. P. Heesterbeek, J. A. J. Metz, On the definition and computation of the basic reproduction ratio $R_0$ in models for infectious diseases in heterogeneous populations, J. Math. Biol., 28 (1990), 365–382. https://doi.org/10.1007/BF00178324 doi: 10.1007/BF00178324
|
| [12] |
S. Li, W. Li, Dynamical behaviors of a stochastic SIVS epidemic model with the Ornstein-Uhlenbeck process and vaccination of newborns, PLoS One, 19 (2024), e0310175. https://doi.org/10.1371/journal.pone.0310175 doi: 10.1371/journal.pone.0310175
|
| [13] |
O. Ogunmiloro, F. O. Abedo, H. A. Kareem, Numerical and stability analysis of the transmission dynamics of the SVIR epidemic model with standard incidence rate, Malays. J. Comput., 4 (2019), 349–361. https://doi.org/10.24191/mjoc.v4i2.5828 doi: 10.24191/mjoc.v4i2.5828
|
| [14] |
S. Hussain, E. N. Madi, H. Khan, S. Etemad, S. Rezapour, T. Sitthiwirattham, et al., Investigation of the stochastic modeling of COVID-19 with environmental noise from the analytical and numerical point of view, Mathematics, 9 (2021), 3122. https://doi.org/10.3390/math9233122 doi: 10.3390/math9233122
|
| [15] | F. Brauer, C. C. Chavez, Mathematical models in epidemiology, 1 Eds., New York: Springer, 2019. https://doi.org/10.1007/978-1-4939-9828-9 |
| [16] |
M. Barman, N. Mishra, Hopf bifurcation in a networked delay SIR epidemic model, J. Math. Anal. Appl., 525 (2023), 127131. https://doi.org/10.1016/j.jmaa.2023.127131 doi: 10.1016/j.jmaa.2023.127131
|
| [17] |
L. Zhu, H. Hu, A stochastic SIR epidemic model with density-dependent birth rate, Adv. Differential Equ., 330 (2015), 1–2. https://doi.org/10.1186/s13662-015-0669-2 doi: 10.1186/s13662-015-0669-2
|
| [18] | L. Allen, An introduction to stochastic epidemic models, 1 Eds., Berlin: Springer-Verlag, 2008, 81–130. https://doi.org/10.1007/978-3-540-78911-6-3 |
| [19] |
H. Alsakaji, Y. El Kahtib, F. Rihan, A. Hashish, A stochastic epidemic model with time delays and unreported cases based on Markovian switching, J. Biol. Dynam., 6 (2024), 234–243. https://doi.org/10.1016/j.jobb.2024.08.002 doi: 10.1016/j.jobb.2024.08.002
|
| [20] |
F. Rao, W. Wang, Z. Li, Stability analysis of an epidemic model with diffusion and stochastic perturbation, Commun. Nonlinear Sci., 7 (2012), 2551–2563. https://doi.org/10.1016/j.cnsns.2011.10.005 doi: 10.1016/j.cnsns.2011.10.005
|
| [21] |
N. Dalal, D. Greenhalgh, X. Mao, A stochastic model of AIDS and condom use, J. Math. Anal. Appl., 325 (2007), 36–53. https://doi.org/10.1016/j.jmaa.2006.01.055 doi: 10.1016/j.jmaa.2006.01.055
|
| [22] |
C. Ji, D. Jiang, Threshold behaviour of a stochastic SIR model, Appl. Math. Model., 38 (2014), 5067–5079. https://doi.org/10.1016/j.apm.2014.03.037 doi: 10.1016/j.apm.2014.03.037
|
| [23] |
D. Higham, An algorithmic introduction to numerical simulation of stochastic differential equations, SIAM Rev., 43 (2001), 525–546. https://doi.org/10.1137/S0036144500378302 doi: 10.1137/S0036144500378302
|





Figures(6)

Shah Hussain, Naveed Iqbal, Elissa Nadia Madi, Thoraya N. Alharthi, Ilyas Khan. Vaccination strategies in a stochastic $ \mathscr{SIVR} $ epidemic model[J]. AIMS Mathematics, 2025, 10(2): 4441-4456. doi: 10.3934/math.2025204







 DownLoad:
DownLoad: