This study introduced a novel $ \mathscr{SVIR} $ epidemic model incorporating environmental white noise to account for stochastic fluctuations in disease transmission. The model was analyzed to determine conditions for disease persistence and extinction, with outcomes linked to the basic reproduction number. A numerical approach was employed to facilitate computational analysis, and simulations were conducted using data from existing literature to generate realistic predictions. The stochastic model was further evaluated against its deterministic counterpart to assess predictive accuracy. The results highlight the significant role of randomness in epidemiological dynamics, providing valuable insights into disease spread and control strategies.
Citation: Shah Hussain, Naveed Iqbal, Elissa Nadia Madi, Mohsen Bakouri, Ilyas Khan, Wei Sin Koh. On the stochastic modeling and forecasting of the $ \mathscr{SVIR} $ epidemic dynamic model under environmental white noise[J]. AIMS Mathematics, 2025, 10(2): 3983-3999. doi: 10.3934/math.2025186
This study introduced a novel $ \mathscr{SVIR} $ epidemic model incorporating environmental white noise to account for stochastic fluctuations in disease transmission. The model was analyzed to determine conditions for disease persistence and extinction, with outcomes linked to the basic reproduction number. A numerical approach was employed to facilitate computational analysis, and simulations were conducted using data from existing literature to generate realistic predictions. The stochastic model was further evaluated against its deterministic counterpart to assess predictive accuracy. The results highlight the significant role of randomness in epidemiological dynamics, providing valuable insights into disease spread and control strategies.

| [1] |
A. Alkhazzan, J. Wang, Y. Nie, H. Khan, J. Alzabut, A stochastic Susceptible Vaccinees Infected Recovered epidemic model with three types of noises, Math. Method. Appl. Sci., 47 (2024), 8748–8770. https://doi.org/10.1002/mma.10042 doi: 10.1002/mma.10042

|
| [2] |
A. Alkhazzan, J. Wang, Y. Nie, H. Khan, J. Alzabut, An effective transport-related SVIR stochastic epidemic model with media coverage and Lévy noise, Chaos Soliton. Fract., 175 (2023), 113953. https://doi.org/10.1016/j.chaos.2023.113953 doi: 10.1016/j.chaos.2023.113953
|
| [3] |
A. Alkhazzan, J. Wang, Y. Nie, K. Hattaf, A new stochastic split-step $\theta$-nonstandard finite difference method for the developed SVIR epidemic model with temporary immunities and general incidence rates, Vaccines, 10 (2022), 1682. https://doi.org/10.3390/vaccines10101682 doi: 10.3390/vaccines10101682
|
| [4] |
H. J. Alsakaji, F. A. Rihan, A. Hashish, Dynamics of a stochastic epidemic model with vaccination and multiple time-delays for COVID-19 in the UAE, Complexity, 2022 (2022), 4247800. https://doi.org/10.1155/2022/4247800 doi: 10.1155/2022/4247800
|
| [5] |
L. J. S. Allen, A primer on stochastic epidemic models: Formulation, numerical simulation, and analysis, Infectious Disease Modelling, 2 (2017), 128–142. https://doi.org/10.1016/j.idm.2017.03.001 doi: 10.1016/j.idm.2017.03.001
|
| [6] |
F. Brauer, Backward bifurcations in simple vaccination/treatment models, J. Biol. Dynam., 5 (2011), 410–418. https://doi.org/10.1080/17513758.2010.510584 doi: 10.1080/17513758.2010.510584
|
| [7] |
A. Gray, D. Greenhalgh, L. Hu, X. Mao, J. Pan, A stochastic differential equation SIS epidemic model, SIAM J. Appl. Math., 71 (2011), 876–902. https://doi.org/10.1137/10081856X doi: 10.1137/10081856X
|
| [8] | G. Rozhnova, A. Nunes, Stochastic effects in a seasonally forced epidemic model, Phys. Rev. E., 82 (2010), 041906. https://doi.org/10.1103/PhysRevE.82.041906 |
| [9] |
P. J. Witbooi, Stability of a stochastic model of an SIR epidemic with vaccination, Acta Biotheor., 65 (2017), 151–165. https://doi.org/10.1007/s10441-017-9308-5 doi: 10.1007/s10441-017-9308-5
|
| [10] |
J. Harianto, T. Suparwati, SVIR epidemic model with non constant population, CAUCHY-Jurnal Matematika Murni dan Aplikasi, 5 (2018), 102–111. https://doi.org/10.18860/ca.v5i3.5511 doi: 10.18860/ca.v5i3.5511
|
| [11] |
Y. Cai, Y. Kang, M. Banerjee, W. Wang, A stochastic epidemic model incorporating media coverage, Commun. Math. Sci., 14 (2016), 893–910. https://doi.org/10.4310/CMS.2016.v14.n4.a1 doi: 10.4310/CMS.2016.v14.n4.a1
|
| [12] |
M. Liu, M. Fan, Permanence of stochastic Lotka-Volterra systems, J. Nonlinear Sci., 27 (2017), 425–452. https://doi.org/10.1007/s00332-016-9337-2 doi: 10.1007/s00332-016-9337-2
|
| [13] |
W. Wang, Y. Cai, Z. Ding, Z. Gui, A stochastic differential equation SIS epidemic model incorporating Ornstein–Uhlenbeck process, Physica A, 509 (2018), 921–936. https://doi.org/10.1016/j.physa.2018.06.099 doi: 10.1016/j.physa.2018.06.099
|
| [14] |
A. Lahrouz, L. Omari, Extinction and stationary distribution of a stochastic SIRS epidemic model with non-linear incidence, Stat. Probabil. Lett., 83 (2013), 960–968. https://doi.org/10.1016/j.spl.2012.12.021 doi: 10.1016/j.spl.2012.12.021
|
| [15] |
X. Zhang, K. Wang, Stochastic model for spread of AIDS driven by Lévy noise, J. Dyn. Diff. Equat., 27 (2015), 215–236. https://doi.org/10.1007/s10884-015-9459-5 doi: 10.1007/s10884-015-9459-5
|
| [16] |
Y. Zhao, D. Jiang, The threshold of a stochastic SIS epidemic model with vaccination, Appl. Math. Comput., 243 (2014), 718–727. https://doi.org/10.1016/j.amc.2014.05.124 doi: 10.1016/j.amc.2014.05.124
|
| [17] |
Q. Yang, D. Jiang, N. Shi, C. Ji, The ergodicity and extinction of stochastically perturbed SIR and SEIR epidemic models with saturated incidence, J. Math. Anal. Appl., 388 (2012), 248–271. https://doi.org/10.1016/j.jmaa.2011.11.072 doi: 10.1016/j.jmaa.2011.11.072
|
| [18] |
C. Ji, D. Jiang, Q. Yang, N. Shi, Dynamics of a multigroup SIR epidemic model with stochastic perturbation, Automatica, 48 (2012), 121–131. https://doi.org/10.1016/j.automatica.2011.09.044 doi: 10.1016/j.automatica.2011.09.044
|
| [19] |
D. Jiang, J. Yu, C. Ji, N. Shi, Asymptotic behavior of global positive solution to a stochastic SIR model, Math. Comput. Model., 54 (2011), 221–232. https://doi.org/10.1016/j.mcm.2011.02.004 doi: 10.1016/j.mcm.2011.02.004
|
| [20] |
R. Rudnicki, Long–time behavior of a stochastic prey–predator model, Stoch. Proc. Appl., 108 (2003), 93–107. https://doi.org/10.1016/S0304-4149(03)00090-5 doi: 10.1016/S0304-4149(03)00090-5
|
| [21] |
F. A. Rihan, H. J. Alsakaji, Dynamics of a stochastic delay differential model for COVID-19 infection with asymptomatic infected and interacting people: Case study in the UAE, Results Phys., 28 (2021), 104658. https://doi.org/10.1016/j.rinp.2021.104658 doi: 10.1016/j.rinp.2021.104658
|
| [22] |
Y. Lin, D. Jiang, T. Liu, Nontrivial periodic solution of a stochastic epidemic model with seasonal variation, Appl. Math. Lett., 45 (2015), 103–107. https://doi.org/10.1016/j.aml.2015.01.021 doi: 10.1016/j.aml.2015.01.021
|
| [23] |
X. Zhang, D. Jiang, A. Alsaedi, T. Hayat, Periodic solutions and stationary distribution of mutualism models in random environments, Physica A, 460 (2016), 270–282. https://doi.org/10.1016/j.physa.2016.05.015 doi: 10.1016/j.physa.2016.05.015
|
| [24] |
Y. Zhao, D. Jiang, The threshold of a stochastic SIS epidemic model with vaccination, Appl. Math. Comput., 243 (2014), 718–727. https://doi.org/10.1016/j.amc.2014.05.124 doi: 10.1016/j.amc.2014.05.124
|
| [25] |
C. Zhu, G. Yin, Asymptotic properties of hybrid diffusion systems, SIAM J. Control Optim., 46 (2007), 1155–1179. https://doi.org/10.1137/060649343 doi: 10.1137/060649343
|
| [26] |
D. J. Higham, An algorithmic introduction to numerical simulation of stochastic differential equations, SIAM Rev., 43 (2001), 525–546. https://doi.org/10.1137/S0036144500378302 doi: 10.1137/S0036144500378302
|
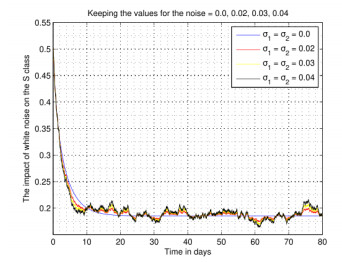



Figures(4)

Shah Hussain, Naveed Iqbal, Elissa Nadia Madi, Mohsen Bakouri, Ilyas Khan, Wei Sin Koh. On the stochastic modeling and forecasting of the $ \mathscr{SVIR} $ epidemic dynamic model under environmental white noise[J]. AIMS Mathematics, 2025, 10(2): 3983-3999. doi: 10.3934/math.2025186







 DownLoad:
DownLoad: