Citation: Kuei Tien Chou. Beyond high carbon society[J]. AIMS Energy, 2016, 4(2): 313-330. doi: 10.3934/energy.2016.2.313

| [1] | German Advisory Council on Global Change (2011) Summary for Policy-Makers World in Transition a Social Contract for Sustainability. German Advisory Council on Global Change, 2. |
| [2] | Chou KT, Liou HM (2012) Analysis on Energy Intensive Industries under Taiwan’s Climate Change Policy. Renew sust energ rev 16: 2631-2642. |
| [3] | IEA (2012) CO2 Emissions from fuel combustion. Paris: International Energy Agency. |
| [4] | Liu CH (2012) How to Implement Sustainable Development of Taiwan? The 9th National Science Technology Conference, Ministry of Science and Technology, Taipei. |
| [5] | Chou KT (2007) Reflexive Discussion on New Risk Governance Paradigm. SOCIETAS: A Journal for Philosophical Study of Public Affairs 22: 179-233. |
| [6] | Chou KT (2013) Governance Innovation of Developmental State under the Globalized Risk Challenges-Burgeoning Civil Knowledge on Risk Policy Supervision in Taiwan. A Journal for Philosophical Study of Public Affairs 44: 65-148. |
| [7] | Chou KT (2014) From Anti-Pollution to Climate Change Risk Movement—Reshaping the Civic Epistemology’ The Seoul Conference with Professor Ulrich Beck: Climate Change and Risk Society: New Trends of Megacity Transformation, Seoul National University, 2014. |
| [8] | Grin J (2006) Reflexive modernization as a governance issue - or: designing and shaping Re- structuration. in: Voß,Jan-Peter; Bauknecht, Dierk; Kemp, René (eds.), Reflexive Governance for Sustainable Development. Cheltenham : EdwardElgar, 54-81. |
| [9] | Beck U (2006) Reflexive Governance: Politics in the Global Risk Society, In J-P. Voß, D.Bauknecht and R. Kemp (Eds.), Reflexive Governance for Sustainable Development, Cheltenham, Edward Elgar. 3-28. |
| [10] |
Hendriks CM, Grin J (2007) Contextualizing Reflexive Governance: the Politics of Dutch Transitions to Sustainability. Routledge, USA. J Environ Pol Plann 9: 333-350. doi: 10.1080/15239080701622790

|
| [11] |
Voβ Jan-Peter, Smith Adrian, Grin John (2009) Designing long time policy: rethinking transition management. Policy Sci 42: 275-302. doi: 10.1007/s11077-009-9103-5
|
| [12] | Wang Jenn-hwan (2012) Long live development studies, City and Planning, 39: 1-18. |
| [13] | Beck U (1993) Die Erfindung des Politischen: Zu einer Theorie reflexiver Modernisierung. Frankfurt am Main: Suhrkamp. Beck U, Giddens A, Lash S (1994). Reflexive modernization: Politics, tradition and aesthetics in the modern social order. Stanford, Calif: Stanford University Press. |
| [14] | Beck U (1997) Subpolitics—Ecology and the Disintegration of Institutional Power, Organization & Environment 10: 52-65. |
| [15] | Nowotny H, Scott P, Gibbons M (2001) The Co-evolution of Society and Science. in: Re-Thinking Science: Knowledge and the public in an age of uncertainty, Cambridge: Polity Press, 30-49. |
| [16] |
Jasanoff S (2003) Technologies of Humility: Citizen Participation in Governing Science. Minerva 41: 223-244. doi: 10.1023/A:1025557512320
|
| [17] |
Stirling A (2007) Risk, precaution and science: towards a more constructive policy debate—Talking point on the precautionary principle. EMBO reports 8: 309-315. doi: 10.1038/sj.embor.7400953
|
| [18] | Delvenne P (2010) Parliamentary technology assessment institutions as indications of reflexive modernization. Society for social studies of science annual meeting with JSSTS. University of Tokyo, Tokyo. |
| [19] | Rip A (2010) Social Robustness and the mode 2 diagnosis. STI studies 6: 71-74. |
| [20] | Irwin A (1995) Citizen science: A study of people, expertise, and sustainable development. London: Routledge. |
| [21] |
Gerold R, Liberatore A (2001) Democratising Expertise and Establishing Scientific Reference Systems. White Paper on Governance, Work Area 1 (Group 1b): Broadening and Enriching the Public Debate on European Matters. Available from: http:// ec.europa.eu/governance/areas/group2/report_en.pdf |
| [22] | Fischer F (1990) ‘Technocracy and Expertise: the Basic Political Question,’ In F. Fischer, Technocracy and the Politics of Expertise, Newbury Park, Calif.: Sage Publications, 13-39. |
| [23] | Jasanoff S (1990) The Fifth Branch: Science Adviser as Policymakers. MA: Harvard University Press. |
| [24] | Hajer Maarten A (1995) The Politics of Environment Discourse: Ecological Modernization and the Policy Process, Oxford: Clarendon Press. |
| [25] | Lash S (1994) Reflexivity and its Doubles: Structure, Aesthetics, Community. In U. Beck, A. Giddens and S. Lash (eds) Reflexive Modernization. Cambridge: Polity Press, 110-173. |
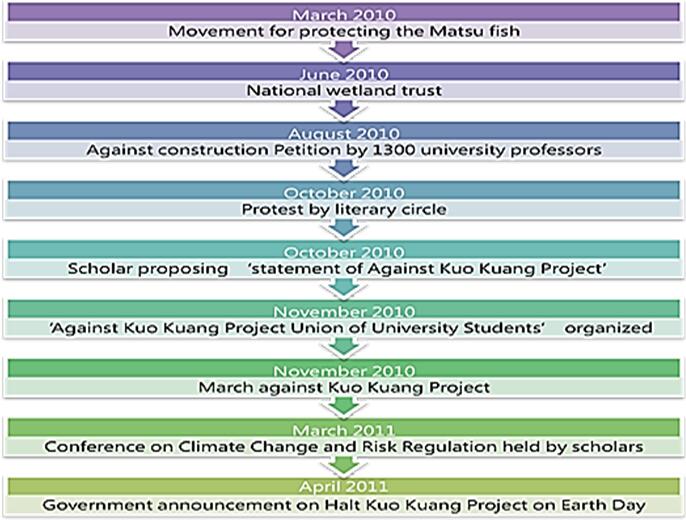
| [26] | Tsai CY (2010) Love and Death of Wet Land of Sea Shore of ChangHwa, Liberty Times. |
| [27] | Lin HJ (2010) Carbon Credit Ecology Service of Wet Land along the Coast of Taiwan, Taiwan Academy of Ecology, 30. |
| [28] | Chen CL (2010) Whey should White Dolphins Make Turns? E-paper of Taiwan Environmenttal Information Center. Available from: http://e-info.org.tw/node/57258. |
| [29] | ChangHwa Environment Protection Union (2010) Real Actions to be Against Kuo Kuang Petrochemical Corp.(the Eighth Naphtha Cracker): National Subscription Protection of White Dolphins. Wet Land, Petrochemical Industry, Island Imagination, Taipei: Route Culture, 232-235. |
| [30] | Literary circle (2010) Mouth of Jhuoshuei River Protected by Literary Circle, Healthy Future for Taiwan, Statement of Petition of Literary Circle against Development of Petrochemical Industry on Mouth of Jhuoshuei River. Available from: http://tepu-cd.blogspot.tw/p/blog-page_02.html. |
| [31] | A-da (2010) Serious situation of Jhuoshuei River, Musical collection of The Village Armed Youth Band. Taichung: GammaMusic. |
| [32] | Wu S (2010) ‘I can only write a poem to you’, Business Weekly, 279. |
| [33] | Chuang FH (2010) ‘Not Just for White Dolphins’, Liberty Times. |
| [34] | Chou KT, et al., (2010) The academic resistance group to Kuo Kuang Petrochemical Corp. (the Eighth Naphtha Cracker) founded in ChangHwa. In: Wu S, Wu MY Editor (Eds.), Wet Land, Petrochemical Industry, Island Imagination, Taipei: Route Culture: 210-2013. |
| [35] | Liu CM (2010) Error of BAU, In: Wu S, Wu MY Editor (Eds.), Wet Land, Petrochemical Industry, Island Imagination. Taipei: Route Culture, 113-115. |
| [36] | Chou KT (2011) Do We Have the Policy of Climate Change? Apple Daily. |
| [37] | The academic resistance group to Kuo Kuang Petrochemical Corp. (the Eighth Naphtha Cracker) founded in ChangHwa (2010) Press conference ‘retort to statements of Ministry of Economic Affairs. Truth (Shi Hua, petrochemical) from academia’. Available from: http://www.coolloud.org.tw/node/55205. |
| [38] | Ministry of Economic Affairs (2010) Industrial development, environmental protection and the public benefit of Ministry of Economic Affairs: policy description to implement Kuo Kuang Petrochemical Corp, Taipei. |
| [39] | Tseng KH (2010) Medical petition against the development of the petrochemical industry, Taiwan Academy of Ecology. Available from: http://www.ecology.org.tw/news/view.php?id=16 |
| [40] | Wynne B, Dressel K (2001) Cultures of uncertainty: Transboundary risks and BSE in Europe. Transboundary Risk Management : 121-154. |
| [41] | Löfstedt R, How can better risk management lead to greater public trust in Canadian institutions: some sobering lessons from Europe, 2003. Available from: http://www.smartregulation.gc.ca. |
| [42] | Chou KT (2015) Cosmopolitan Approach of Trans-boundary Risk Governance in East Asia, World Congress of Risk Analysis, Singapore. |
| [43] |
Chou KT (2013) The Public Perception of Climate Change in Taiwan in Paradigm Shift. Energy Policy 61: 1252-1260. doi: 10.1016/j.enpol.2013.06.016
|
| [44] |
Beck U (2015) Emacipatory Catastrophism: What does it mean to Climate Change and Risk Society? Current Sociology 63: 75-88. doi: 10.1177/0011392114559951
|
Figures(1)

Kuei Tien Chou. Beyond high carbon society[J]. AIMS Energy, 2016, 4(2): 313-330. doi: 10.3934/energy.2016.2.313







 DownLoad:
DownLoad: