Citation: Juan Wang, Chunyang Qin, Yuming Chen, Xia Wang. Hopf bifurcation in a CTL-inclusive HIV-1 infection model with two time delays[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 2587-2612. doi: 10.3934/mbe.2019130

| [1] | A. L. Cunningham, H. Donaghy, A. N. Harman, et al., Manipulation of dendritic cell function by viruses, Curr. Opin. Microbiol., 13 (2010), 524–529. |
| [2] | A. S. Perelson and R. M. Ribeiro, Modeling the within-host dynamics of HIV infection, BMC Biology, 11 (2013), 96. |
| [3] | R. M. Riberio and A. S. Perelson, The analysis of HIV dynamics using mathematical modeling, in AIDS and other manisfestation of HIV infection, (Edited by G.P. Wormser), San Diego, Elsevier, (2004), 905–912. |
| [4] | P. De Leenheer and H. L. Smith, Virus dynamics: a global analysis, SIAM J. Appl. Math., 63 (2003), 1313–1327. |
| [5] | A. M. Elaiw and S. A. Azoz, Global properties of a class of HIV infection models with Beddington-DeAngelis functional response, Math. Methods Appl. Sci., 36 (2013), 383–394. |
| [6] | X. Lai and X. Zou, Modeling cell-to-cell spread of HIV-1 with logistic target cell growth, J. Math. Anal. Appl., 426 (2015), 563–584. |
| [7] | K. A. Pawelek, S. Liu, F. Pahlevani, et al., A model of HIV-1 infection with two time delays: mathematical analysis and comparison with patient data, Math. Biosci., 235 (2012), 98–109. |
| [8] | A. S. Perelson, D. E. Kirschner and R. de Boer, Dynamics of HIV infection of CD4+ T cells, Math. Biosci., 114 (1993), 81–125. |
| [9] | A. S. Perelson and P. W. Nelson, Mathematical analysis of HIV-1 dynamics in vivo, SIAM Rev., 41 (1999), 3–44. |
| [10] | P. K. Srivastava, M. Banerjee and P. Chandra, A primary infection model for HIV and immune response with two discrete time delays, Differ. Equ. Dyn. Syst., 18 (2010), 385–399. |
| [11] | P. K. Srivastava and P. Chandra, Hopf bifurcation and periodic solutions in a dynamical model for HIV and immune response, Differ. Equ. Dyn. Syst., 16 (2008), 77–100. |
| [12] | H. Wang, R. Xu, Z. Wang, et al., Global dynamics of a class of HIV-1 infection models with latently infected cells, Nonlinear Anal. Model. Control, 20 (2015), 21–37. |
| [13] | X. Wang, Y. Lou and X. Song, Age-structured within-host HIV dynamics with multiple target cells, Stud. Appl. Math., 138 (2017), 43–76. |
| [14] | X. Wang, G. Mink, D. Lin, et al., Influence of raltegravir intensification on viral load and 2-LTR dynamics in HIV patients on suppressive antiretroviral therapy, J. Theor. Biol., 416 (2017), 16–27. |
| [15] | X. Wang, X. Song, S. Tang, et al., Dynamics of an HIV model with multiple infection stages and treatment with different drug classes, Bull. Math. Biol., 78 (2016), 322–349. |
| [16] | Y. Wang, Y. Zhou, F. Brauer, et al., Viral dynamics model with CTL immune response incorporating antiretroviral therapy, J. Math. Biol., 67 (2013), 901–934. |
| [17] | N. M. Dixit, M. Markowitz, D. D. Ho, et al., Estimates of intracellular delay and average drug efficacy from viral load data of HIV-infected individuals under antiretroviral therapy, Antivir. Ther., 9 (2004), 237–246. |
| [18] | A. V. M. Herz, S. Bonhoeffer, R. M. Anderson, et al., Viral dynamics in vivo: limitations on estimates of intracellular delay and virus decay, Proc. Nat. Acad. Sci. USA, 93 (1996), 7247– 7251. |
| [19] | K. Allali, S. Harroudi and D. F. M. Torres, Analysis and optimal control of an intracellular delayed HIV model with CTL immune response, Math. Comput. Sci., 12 (2018), 111–127. |
| [20] | M. S. Ciupe, B. L. Bivort, D. M. Bortz, et al., Estimating kinetic parameters from HIV primary infection data through the eyes of three different mathematical models, Math. Biosci., 200 (2006), 1–27. |
| [21] | R. V. Culshaw and S. Ruan, A delay-differential equation model of HIV infection of CD4+ T cells, ath. Biosci., 165 (2000), 27–39. |
| [22] | R. V. Culshaw, S. Ruan and G. Webb, A mathematical model of cell-to-cell HIV-1 that include a time delay, J. Math. Biol., 46 (2003), 425–444. |
| [23] | B. Li, Y. Chen, X. Lu, et al., A delayed HIV-1 model with virus waning term, Math. Biosci. Eng., 13 (2016), 135–157. |
| [24] | Y. Liu and C. Wu, Global dynamics for an HIV infection model with Crowley-Martin functional response and two distributed delays, J. Syst. Sci. Complex., 31 (2018), 385–395. |
| [25] | Y. Wang, Y. Zhou, J. Wu, et al., Oscillatory viral dynamics in a delayed HIV pathogenesis model, Math. Biosci., 219 (2009), 104–112. |
| [26] | H. Zhu and X. Zou, Dynamics of a HIV-1 Infection model with cell-mediated immune response and intracellular delay, Discrete Contin. Dyn. Syst. Ser. B, 12 (2009), 511–524. |
| [27] | N. Tarfulea, A. Blink, E. Nelson, et al., A CTL-inclusive mathematical model for antiretroviral treatment of HIV infection, Int. J. Biomath., 4 (2011), 1–22. |
| [28] | P. W. Nelson, J. D. Murray and A. S. Perelson, A model of HIV-1 pathogenesis that Includes an intracellular delay, Math. Biosci., 163 (2000), 201–215. |
| [29] | J. K. Hale and S. Verduyn Lunel, Introduction to Functional Differential Equations, Springer- Verlag, New York, 1993. |
| [30] | H. R. Thieme, Spectral bound and reproduction number for infinite-dimensional population structure and time heterogeneity, SIAM J. Appl. Math., 70 (2009), 188–211. |
| [31] | J. LaSalle and S. Lefschetz, Stability by Liapunov's Direct Method, with Applications, Academic Press, New York, 1961. |
| [32] | E. Beretta and Y. Kuang, Geometric stability switch criteria in delay differential systems with delay dependent parameters, SIAM J. Math. Anal., 33 (2002), 1144–1165. |
| [33] | X. Yan andW. Li, Stability and bifurcation in a simplified four-neural BAM network with multiple delays, Discrete Dyn. Nat. Soc., 2006 (2006), 1–29. |
| [34] | M. Y. Li and H. Shu, Multiple stable periodic oscillations in a mathematical model of CTL response to HTLV-I infection, Bull. Math. Biol., 73 (2011), 1774–1793. |
| [35] | M. Y. Li, X. Lin and H. Wang, Global Hopf branches in a delayed model for immune response to HTLV-1 infections: coexistence of multiple limit cycles, Can. Appl. Math. Q., 20 (2012), 39–50. |
| [36] | A. Debadatta and B. Nandadulal, Analysis and computation of multi-pathways and multi-delays HIV-1 infection model, Appl. Math. Model., 54 (2018), 517–536. |
| [37] | B. M. Adams, H. T. Banks, M. Davidian, et al., HIV dynamics: modeling, data analysis, and optimal treatment protocols, J. Comput. Appl. Math., 184 (2005), 10–49. |
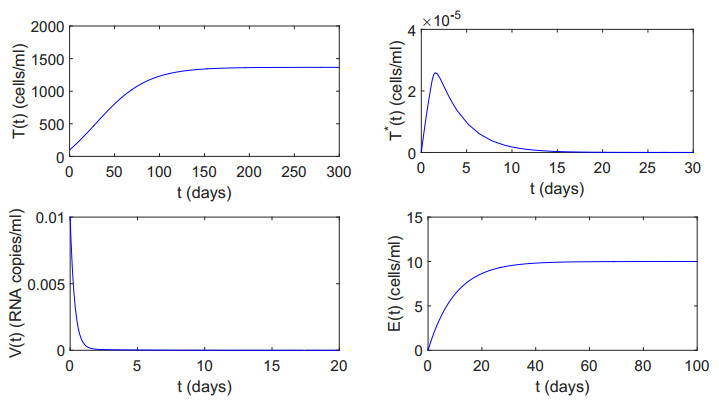
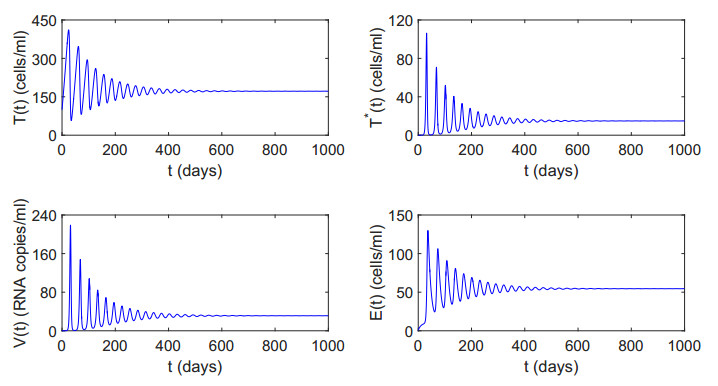
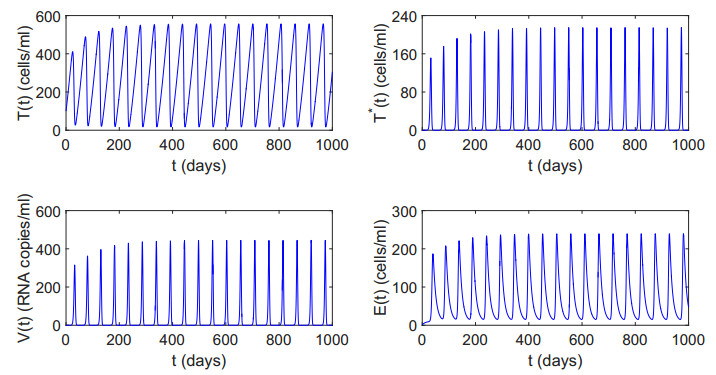
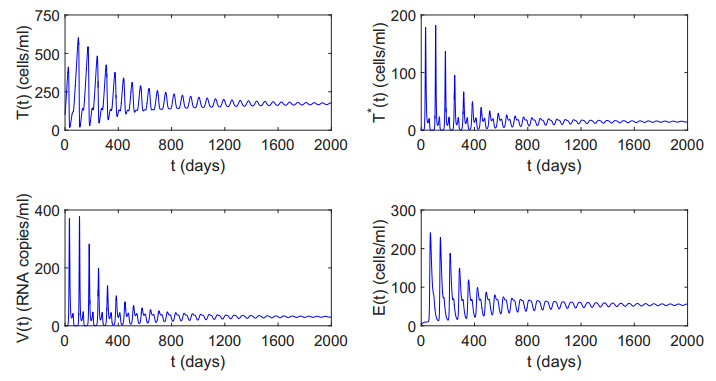
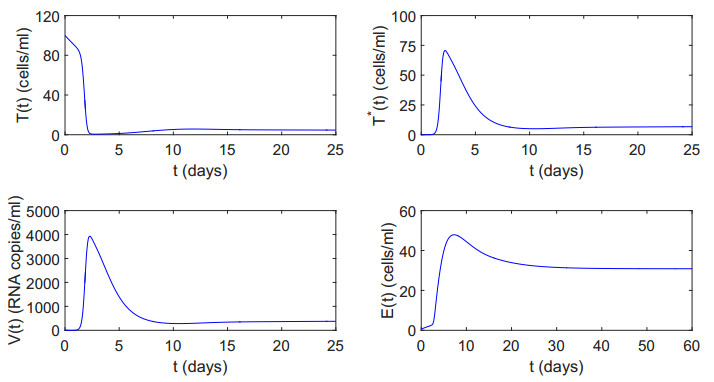
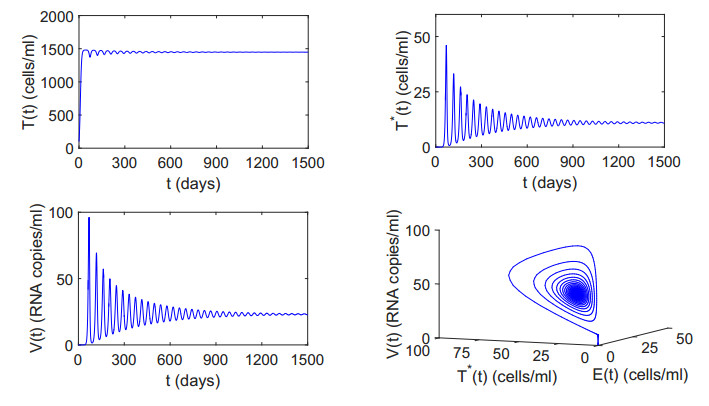
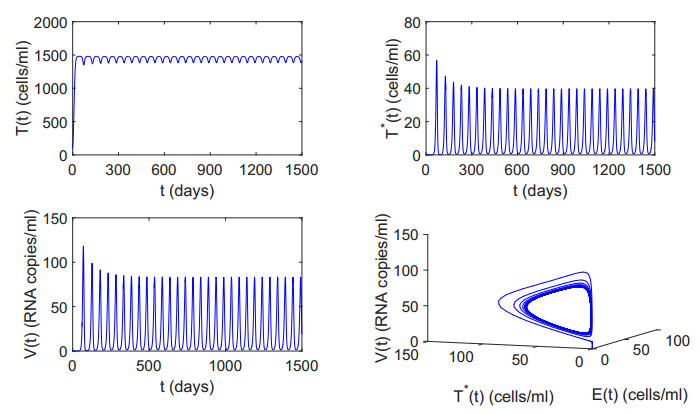
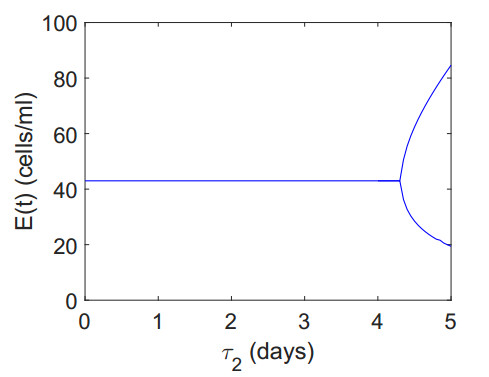
Figures(12) / Tables(2)

Juan Wang, Chunyang Qin, Yuming Chen, Xia Wang. Hopf bifurcation in a CTL-inclusive HIV-1 infection model with two time delays[J]. Mathematical Biosciences and Engineering, 2019, 16(4): 2587-2612. doi: 10.3934/mbe.2019130







 DownLoad:
DownLoad: